

Synaptic variance and action potential firing of cerebellar output neurons during motor learning in larval zebrafish
- PMID: 37421952
- PMCID: PMC10527510
- DOI: 10.1016/j.cub.2023.06.045
Synaptic variance and action potential firing of cerebellar output neurons during motor learning in larval zebrafish
Abstract
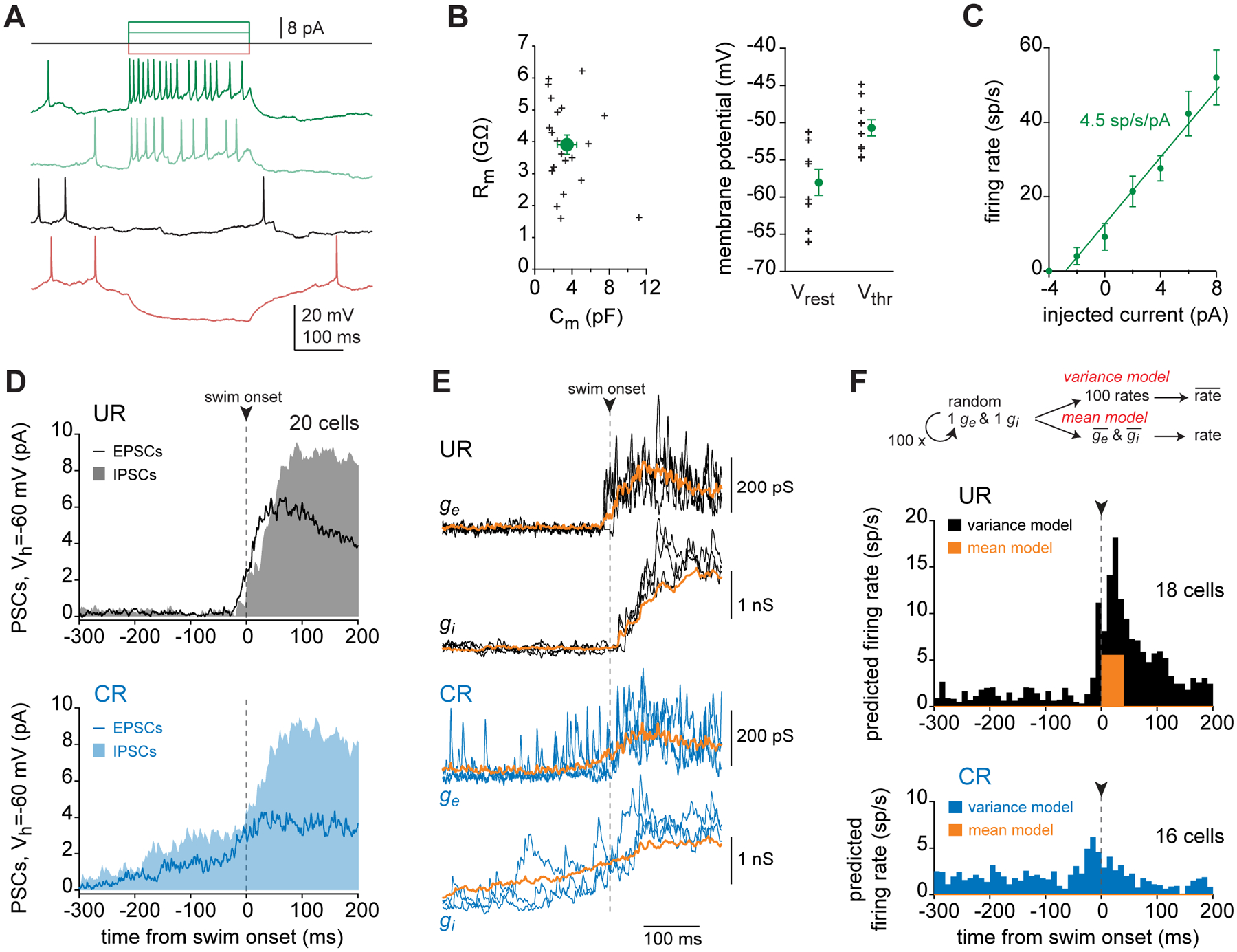
The cerebellum regulates both reflexive and acquired movements. Here, by recording voltage-clamped synaptic currents and spiking in cerebellar output (eurydendroid) neurons in immobilized larval zebrafish, we investigated synaptic integration during reflexive movements and throughout associative motor learning. Spiking coincides with the onset of reflexive fictive swimming but precedes learned swimming, suggesting that eurydendroid signals may facilitate the initiation of acquired movements. Although firing rates increase during swimming, mean synaptic inhibition greatly exceeds mean excitation, indicating that learned responses cannot result solely from changes in synaptic weight or upstream excitability that favor excitation. Estimates of spike threshold crossings based on measurements of intrinsic properties and the time course of synaptic currents demonstrate that noisy excitation can transiently outweigh noisy inhibition enough to increase firing rates at swimming onset. Thus, the millisecond-scale variance of synaptic currents can regulate cerebellar output, and the emergence of learned cerebellar behaviors may involve a time-based code.
Keywords: EPSC; IPSC; cerebellum; conditioning; eurydendroid; fictive swimming; spike timing.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





Comment in
-
Cerebellar nuclei: Associative motor learning in zebrafish.Curr Biol. 2023 Aug 21;33(16):R867-R870. doi: 10.1016/j.cub.2023.07.018. Curr Biol. 2023. PMID: 37607484
Similar articles
-
Integration of Swimming-Related Synaptic Excitation and Inhibition by olig2+ Eurydendroid Neurons in Larval Zebrafish Cerebellum.J Neurosci. 2020 Apr 8;40(15):3063-3074. doi: 10.1523/JNEUROSCI.2322-19.2020. Epub 2020 Mar 5. J Neurosci. 2020. PMID: 32139583 Free PMC article.
-
Distinct responses of Purkinje neurons and roles of simple spikes during associative motor learning in larval zebrafish.Elife. 2017 May 25;6:e22537. doi: 10.7554/eLife.22537. Elife. 2017. PMID: 28541889 Free PMC article.
-
Decreased intrinsic excitability of cerebellar Purkinje cells following optokinetic learning in mice.Mol Brain. 2020 Oct 7;13(1):136. doi: 10.1186/s13041-020-00678-2. Mol Brain. 2020. PMID: 33028375 Free PMC article.
-
The neuronal code(s) of the cerebellum.J Neurosci. 2013 Nov 6;33(45):17603-9. doi: 10.1523/JNEUROSCI.2759-13.2013. J Neurosci. 2013. PMID: 24198351 Free PMC article. Review.
-
What features of limb movements are encoded in the discharge of cerebellar neurons?Cerebellum. 2011 Dec;10(4):683-93. doi: 10.1007/s12311-010-0243-0. Cerebellum. 2011. PMID: 21203875 Free PMC article. Review.
Cited by
-
Cerebellar Purkinje Cells Control Posture in Larval Zebrafish (Danio rerio).bioRxiv [Preprint]. 2024 Nov 9:2023.09.12.557469. doi: 10.1101/2023.09.12.557469. bioRxiv. 2024. Update in: Elife. 2025 Apr 24;13:RP97614. doi: 10.7554/eLife.97614. PMID: 37745506 Free PMC article. Updated. Preprint.
-
Cerebellar Purkinje cells control posture in larval zebrafish (Danio rerio).Elife. 2025 Apr 24;13:RP97614. doi: 10.7554/eLife.97614. Elife. 2025. PMID: 40272244 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
