

Stress and Liver Fibrogenesis: Understanding the Role and Regulation of Stress Response Pathways in Hepatic Stellate Cells
- PMID: 37422148
- PMCID: PMC10548279
- DOI: 10.1016/j.ajpath.2023.06.006
Stress and Liver Fibrogenesis: Understanding the Role and Regulation of Stress Response Pathways in Hepatic Stellate Cells
Abstract
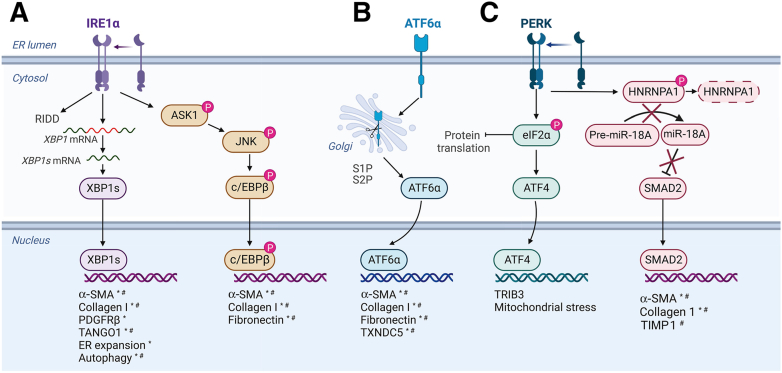
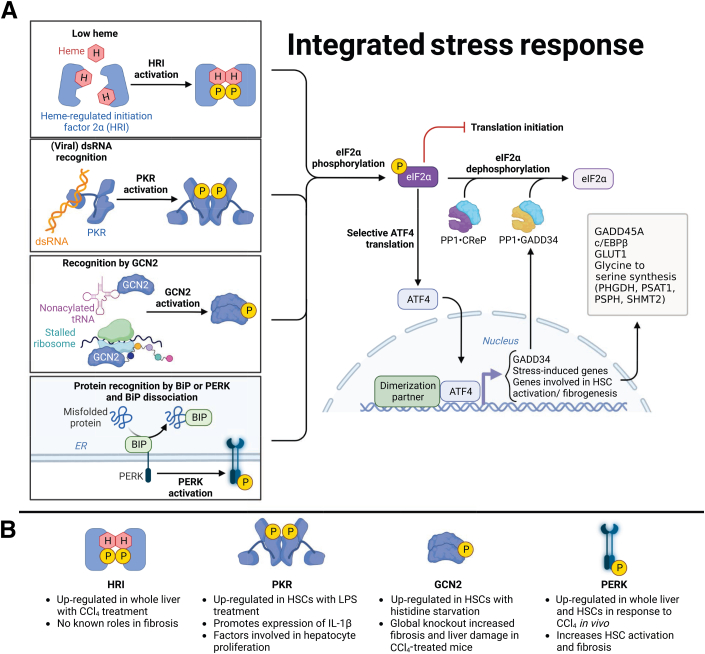
Stress response pathways are crucial for cells to adapt to physiological and pathologic conditions. Increased transcription and translation in response to stimuli place a strain on the cell, necessitating increased amino acid supply, protein production and folding, and disposal of misfolded proteins. Stress response pathways, such as the unfolded protein response (UPR) and the integrated stress response (ISR), allow cells to adapt to stress and restore homeostasis; however, their role and regulation in pathologic conditions, such as hepatic fibrogenesis, are unclear. Liver injury promotes fibrogenesis through activation of hepatic stellate cells (HSCs), which produce and secrete fibrogenic proteins to promote tissue repair. This process is exacerbated in chronic liver disease, leading to fibrosis and, if unchecked, cirrhosis. Fibrogenic HSCs exhibit activation of both the UPR and ISR, due in part to increased transcriptional and translational demands, and these stress responses play important roles in fibrogenesis. Targeting these pathways to limit fibrogenesis or promote HSC apoptosis is a potential antifibrotic strategy, but it is limited by our lack of mechanistic understanding of how the UPR and ISR regulate HSC activation and fibrogenesis. This article explores the role of the UPR and ISR in the progression of fibrogenesis, and highlights areas that require further investigation to better understand how the UPR and ISR can be targeted to limit hepatic fibrosis progression.
Copyright © 2023 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
The Cellular, Molecular, and Pathologic Consequences of Stress on the Liver.Am J Pathol. 2023 Oct;193(10):1353-1354. doi: 10.1016/j.ajpath.2023.07.003. Epub 2023 Aug 4. Am J Pathol. 2023. PMID: 37544504 Free PMC article. No abstract available.
Similar articles
-
Coordinated signaling of activating transcription factor 6α and inositol-requiring enzyme 1α regulates hepatic stellate cell-mediated fibrogenesis in mice.Am J Physiol Gastrointest Liver Physiol. 2021 May 1;320(5):G864-G879. doi: 10.1152/ajpgi.00453.2020. Epub 2021 Mar 17. Am J Physiol Gastrointest Liver Physiol. 2021. PMID: 33728997 Free PMC article.
-
The unfolded protein response mediates fibrogenesis and collagen I secretion through regulating TANGO1 in mice.Hepatology. 2017 Mar;65(3):983-998. doi: 10.1002/hep.28921. Epub 2016 Dec 31. Hepatology. 2017. PMID: 28039913 Free PMC article.
-
Liver Fibrosis: From Basic Science towards Clinical Progress, Focusing on the Central Role of Hepatic Stellate Cells.Int J Mol Sci. 2024 Jul 18;25(14):7873. doi: 10.3390/ijms25147873. Int J Mol Sci. 2024. PMID: 39063116 Free PMC article. Review.
-
The XBP1 Arm of the Unfolded Protein Response Induces Fibrogenic Activity in Hepatic Stellate Cells Through Autophagy.Sci Rep. 2016 Dec 20;6:39342. doi: 10.1038/srep39342. Sci Rep. 2016. PMID: 27996033 Free PMC article.
-
Metabolism of hepatic stellate cells in chronic liver diseases: emerging molecular and therapeutic interventions.Theranostics. 2025 Jan 2;15(5):1715-1740. doi: 10.7150/thno.106597. eCollection 2025. Theranostics. 2025. PMID: 39897543 Free PMC article. Review.
Cited by
-
Arctigenin Attenuates Hepatic Stellate Cell Activation via Endoplasmic Reticulum-Associated Degradation (ERAD)-Mediated Restoration of Lipid Homeostasis.J Agric Food Chem. 2025 Jun 4;73(22):13918-13933. doi: 10.1021/acs.jafc.5c01366. Epub 2025 May 25. J Agric Food Chem. 2025. PMID: 40415275 Free PMC article.
-
The Cellular, Molecular, and Pathologic Consequences of Stress on the Liver.Am J Pathol. 2023 Oct;193(10):1353-1354. doi: 10.1016/j.ajpath.2023.07.003. Epub 2023 Aug 4. Am J Pathol. 2023. PMID: 37544504 Free PMC article. No abstract available.
References
-
- Scaglione S., Kliethermes S., Cao G., Shoham D., Durazo R., Luke A., Volk M.L. The epidemiology of cirrhosis in the United States: a population-based study. J Clin Gastroenterol. 2015;49:690–696. - PubMed
-
- Asrani S.K., Devarbhavi H., Eaton J., Kamath P.S. Burden of liver diseases in the world. J Hepatol. 2019;70:151–171. - PubMed
-
- Tsuchida T., Friedman S.L. Mechanisms of hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol. 2017;14:397–411. - PubMed
-
- Ezhilarasan D., Sokal E., Najimi M. Hepatic fibrosis: it is time to go with hepatic stellate cell-specific therapeutic targets. Hepatobiliary Pancreat Dis Int. 2018;17:192–197. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
