

doi: 10.1093/plphys/kiad301.
Stop and go signals at the stigma-pollen interface of the Brassicaceae
Affiliations
- PMID: 37423711
- PMCID: PMC10517188
- DOI: 10.1093/plphys/kiad301
Item in Clipboard
Stop and go signals at the stigma-pollen interface of the Brassicaceae
Plant Physiol.
.
Erratum in
-
Correction to: Stop and go signals at the stigma-pollen interface of the Brassicaceae.Plant Physiol. 2024 Jul 31;195(4):3138. doi: 10.1093/plphys/kiae242. Plant Physiol. 2024. PMID: 38717872 Free PMC article. No abstract available.
No abstract available
Conflict of interest statement
Conflict of interest statement. None declared.
Figures
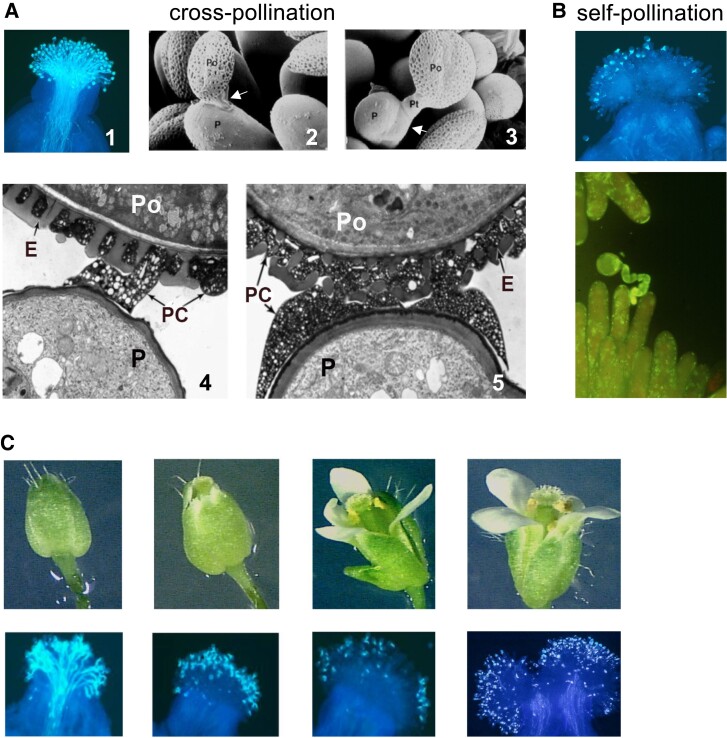
Cross- and self-pollination in crucifers. A) Pollination with cross-pollen. (1) Profuse pollen-tube growth as visualized by light microscopy. The images were generated by fixing pollinated stigmas, treating them with decolorized aniline blue which stains the callose in pollen tube walls, and visualization by UV fluorescence microcopy. (2 and 3) The pollen–papilla interaction as visualized by scanning electron microscopy. Arrows in (2) and (3) point to the attachment foot and to a pollen tube growing within the papilla cell wall, respectively. (4 and 5) Establishment of the attachment foot at the pollen–papilla interface as visualized by TEM. The images were taken at 5-min intervals during establishment of the attachment foot. B) Pollination with self-pollen. Most pollen grains are inhibited at the stigma surface due to failure of pollen hydration and germination (top), but in the few cases where the incompatible grain germinates, its tube fails to grow into the papilla cell wall (bottom). The light microscopy images were generated as in A), except that a different filter was used for UV fluorescence microscopy in the lower image. C) Regulation of self-incompatibility as a function of stigma maturation in developing floral buds. A developmental series of floral buds (top) and images of the self-pollinated stigmas from buds at similar stages of development (bottom) are shown. The stigmas are initially able to accept self-pollen shortly before flower opening. The light microscopy images in A and B) (top image) are reprinted from Nasrallah (2017), and the image in B) (bottom) is modified from Nasrallah (1997). The TEM images are reprinted from Kandasamy et al. (1994), and the floral-bud images are reprinted from Nasrallah and Nasrallah (2014). P, papilla; Po, pollen grain; PC, pollen coat; E, exine.
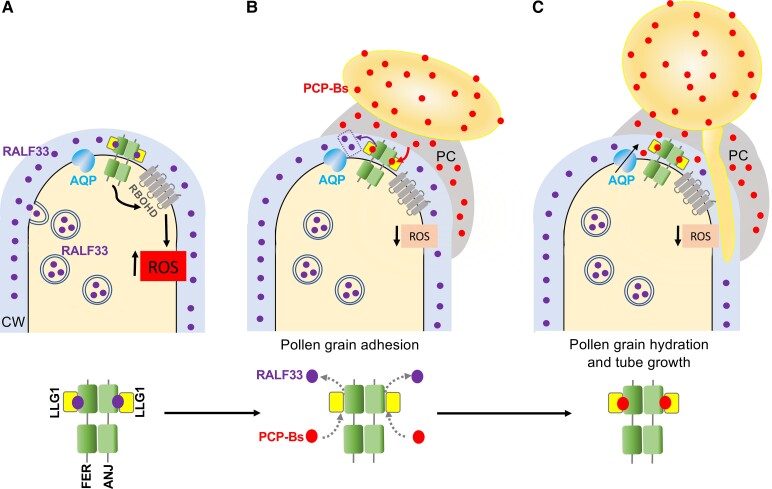
Proposed mechanism for hydration of compatible pollen grains based on the regulation of ROS levels by the FER/ANJ/LLG1 receptor complex. The figure shows a simplified diagram of this process in the papilla (upper diagrams) and a magnified view of the interaction of FER/ANJ/LLG1 with its ligands (lower diagrams). A) In the unpollinated stigma papilla of A. thaliana, the RALF33 peptide is secreted to the cell wall (CW) where it binds to the FER/ANJ/LLG1 complex and activates ROS production by RBOHD, thus maintaining high ROS levels. B) In a compatible pollination, pollen coat (PC) PCP-B peptides released from the pollen grain as the PC flows onto the papilla cell wall outcompete RALF33 for binding to the FER/ANJ/LLF1 complex causing reduced receptor signaling and low ROS levels. C) The reduced ROS levels caused by the compatible pollen–papilla interaction promote water transfer from papilla to pollen grain likely via aquaporin (AQP) channels, thus allowing pollen grain hydration, germination, and tube growth within the papilla cell wall. Not shown are intermediates of the FER/ANJ/LLG1-to-RBOH pathway, the reported increase in nitric oxide induced by compatible pollen with subsequent nitrosation of FER, and the various other cellular events known to be triggered by compatible pollination which are thought to promote water release from the papilla (see text). Note that while the displacement of RALF33 by PCP-Bs must occur in the region of the papilla that subtends the pollen grain as shown in B and C), it is not known how localized the effect of reduced FER/ANJ/LLG1 signaling is within the papilla.
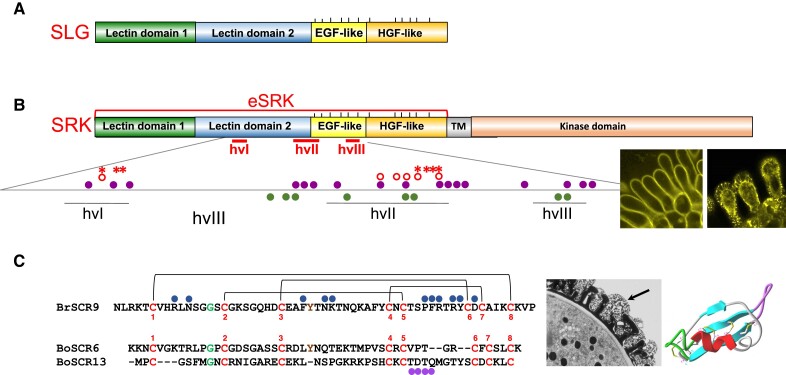
Structure of S locus-encoded proteins. A) Structure of the SLG protein. The diagram shows the modular organization that characterizes the S-domain in members of the SD-RLK/RLP family. The S-domain consists of 2 N-terminal lectin domains, an epidermal growth factor (EGF)-like domain, and a hepatocyte growth factor (HGF)-like domain. The vertical lines above the diagram mark the twelve cysteine residues that are conserved in S-domain proteins. The SLG gene occurs in most S haplotypes from Brassica and Raphanus spp. but not in S haplotypes from Arabidopsis and Capsella spp. B) Structure of the SRK protein. The protein consists of an extracellular S domain (eSRK), a transmembrane domain (TM), and a kinase domain. The location of the 3 hypervariable (hv) regions that contain the SI specificity determinants and the amino acids that form the SRK-SCR binding pocket are shown. Below the diagram is an enlarged view of the hv-containing segment that shows the location of amino-acid residues found to be critical for SRK function. The amino-acid residues essential for in-planta activation of SI in 2 SRK variants are shown by open circles for one variant and asterisks for the second variant (Boggs et al. 2009c). The enlargement also shows the amino acids that form contact points with SCR9 (filled circles above the line) and those involved in eSRK9 homodimerization (filled circles below the line) as determined by the 3D crystal structure of the eSRK9-SCR9 tetrameric complex. Note that residues in hvIII were not tested for activity in planta. To the right of the diagram are confocal images of an AlSRK20-YFP fusion protein which confers SI in A. thaliana C24 stigmas. The images show that the AlSRK20-YFP signal is primarily localized at the periphery of papillae (left) and in the plasma membrane and hechtian strands in plasmolyzed papillae (right). C) Structure and extreme diversity of the SCR protein. Sequence alignment of B. rapa SCR9 (BrSCR9) and B. oleracea SCR6 (BoSCR6) and SCR13 (BoSCR13). The conserved cysteines are shown joined by 4 di-sulfide bridges (brackets above the BrSCR9 sequence). The sequences show the conserved glycine residue and the aromatic residue between C3 and C4 which is conserved in most SCRs including BrSCR9 and BoSCR6 but is missing in BoSCR13. BrSCR9 amino acids that form contact points with eSRK9 in the crystal structure of the B. rapa eSRK9-SCR9 complex are indicated by filled circles above the BsSCR9 sequence. The TDTQ amino acid sequence in BoSCR13 which determines S13 specificity is indicated by filled circles below the BoSCR6 and BoSCR13 sequence alignment. Note that the specificity determinants in SCR6 could not be identified either because they are not located exclusively in the same region as SCR13 determinants or because residues outside this region affect the conformation of the SRK binding pocket. To the right of the alignments is a TEM image of the pollen grain with the arrow pointing to the pollen coat and a 3D model of the BoSCR13 protein showing a structure composed of an α-helix and a triple-stranded antiparallel β-sheet forming a defensin-like cystine-stabilized αβ structure. The SCR13 specificity-determining TDTQ sequence is located in the unstructured loop that projects from the core of the 3D model. The confocal images in B) are reprinted from Tantikanjana and Nasrallah (2015). The 3D model of BoSCR13 in C) is reprinted from Chookajorn et al. (2004).
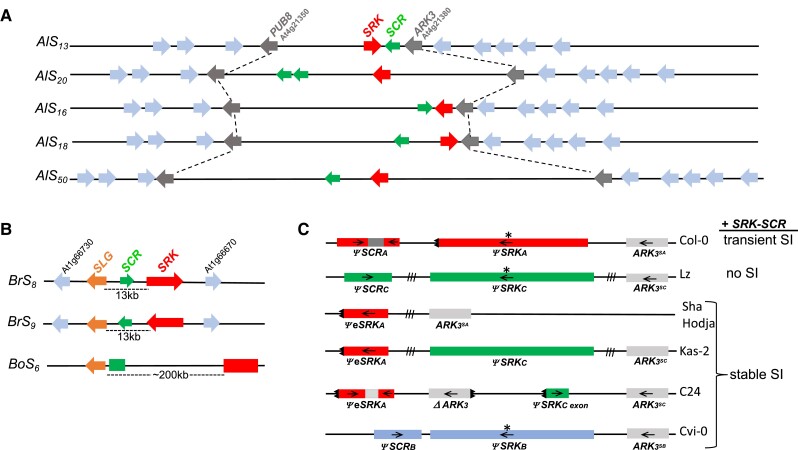
Structural heteromorphism of S haplotypes. The figure shows the rearranged gene order of representative S haplotypes from Arabidopsis and Brassica spp. A) S haplotypes from self-incompatible A. lyrata; B) S haplotypes from self-incompatible B. rapa (Br) and B. oleracea (Bo); C) nonfunctional S haplotypes from several A. thaliana accessions. The arrows indicate the 5′→3′ orientation of each gene, while boxes with no arrows indicate that gene orientation is not known. The diagrams are not drawn to scale. A and B) The Brassica S haplotypes contain an SLG gene while A. lyrata haplotypes do not. In Arabidopsis spp. and Capsella spp. (not shown), the S haplotype is located in a region that corresponds to the segment flanked by the PUB8 (At4g21350) and ARK3 (At4g21380) genes on chromosome 4 of A. thaliana. The Brassica S locus was translocated to a genomic location that corresponds to a region on chromosome 1 of A. thaliana (Conner et al. 1998). The S haplotypes differ in their overall size and with respect to the distance between SRK and SCR and the orientation of these genes relative to each other and to flanking genes. This rearranged gene order can be contrasted with the collinearity of genes that flank the S haplotype in Arabidopsis spp. C) The organization of the nonfunctional S haplotypes in several A. thaliana accessions are shown along with the pollination phenotype conferred by the AlSRK20-AlSCR20 transgenes in each accession. Sequences from the 3 S haplogroups, SA, SB, and SC, which have been retained in the species are shown. The S haplotypes carry inactivating substitutions (asterisks), deletions, inversions, or inter-haplogroup recombination events. AlSRK20-AlSCR20 transformants of the C24, Sha, Hodja, Kas-2, C24, and Cvi-0 accessions exhibit complete reversion to SI, indicating that inactivation of the S locus caused loss of SI in these accessions. Col-0[AlSRK20-AlSCR20] transformants exhibit transient SI due to suboptimal levels of SRK at later stages of sigma development, while Lz[AlSRK20-AlSCR20] plants do not express SI at any stage of stigma development despite expressing adequate levels of AlSRK20 and AlSCR20 transcripts, possibly due to the presence of mutation(s) in the SI pollen rejection pathway. Arrows show the 5′→3′ orientation of the sequences and teeth marks indicate 5′ and 3′ gene truncations. Hatch marks between the genes or gene fragments indicate the segments for which length and sequence content are not known. The maps were drawn according to Kusaba et al. (2001) for AlS13 and AlS20 (previously referred to as AlSa and AlSb, respectively), Guo et al. (2011) for AlS16 and AlS50, and Goubet et al. (2012) for AlS18. C) was modified from Nasrallah (2017).
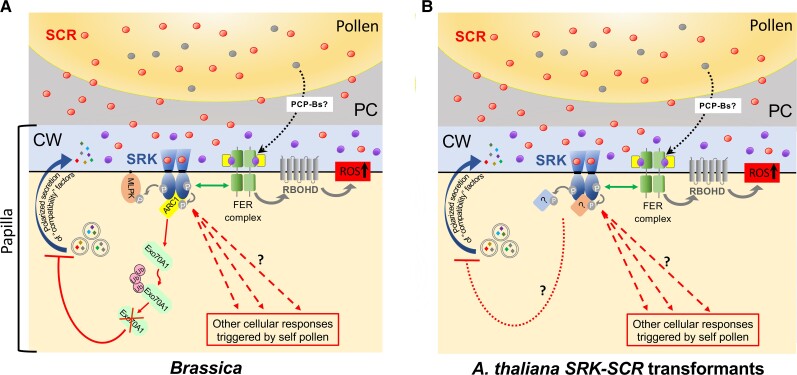
Molecular events underlying self-incompatible interactions at the papilla–pollen grain interface in Brassica and transgenic A. thaliana SRK-SCR plants. Naturally occurring self-incompatible plants such as Brassica plants are heterozygous at the S locus; thus, a stigmatic papilla expresses 2 different SRK variants and a pollen grain generally carries 2 SCR variants (except in the case of dominant-recessive SCR interactions as described in the text). For simplicity, only 1 SRK variant and its cognate SCR are shown in the Brassica diagram. On the other hand, transgenic A. thaliana SRK-SCR plants, which are typically designed to express one SI specificity, are hemizygous (primary transformants) or homozygous (transgenic progenies) for 1 SRK-SCR gene pair. A and B) The diagrams depict the interaction of SRK with its cognate SCR in Brassica and self-incompatible A. thaliana SRK-SCR transgenic plants, the resulting autophosphorylation of the receptor, subsequent phosphorylation of downstream substrates, and proposed SRK-mediated signaling pathways that lead to inhibition of self-pollen. Inhibition of polarized secretion of “compatibility” factors (curved arrow) and maintenance of high ROS levels by interaction (double-headed arrow) between SRK and the FER-ANJ-LLG1 complex (FER complex bound to the RALF33 peptide) are shown as major outcomes of SRK-mediated signaling. Several hypothetical pathways which together produce the various cellular responses triggered by self-pollen (see text) are shown as operating in parallel (dashed arrows with question marks), but intersecting pathways are also possible. How PCP-Bs impact the proposed SRK-mediated pathways is not known. A) In Brassica, MLPK, ARC1, and the ARC1-Exo70A1 pathway leading to degradation of ubiquinated Exo70A1 and inhibition of polarized secretion (blunt curved arrow) are shown. B) In A. thaliana, the downstream targets of SRK are not known (boxes with question marks) and the unknown mechanism by which polarized secretion is inhibited is indicated by the dashed blunt curved arrow. CW, cell wall; PC, pollen coat.
References
-
- Bateman AJ. Self-incompatibility systems in angiosperms. III. Cruciferae. Heredity. 1955:9(1):53–68. 10.1038/hdy.1955.2 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
