

This is a preprint.
Toxic anti-phage defense proteins inhibited by intragenic antitoxin proteins
- PMID: 37425788
- PMCID: PMC10327210
- DOI: 10.1101/2023.05.02.539157
Toxic anti-phage defense proteins inhibited by intragenic antitoxin proteins
Update in
-
Toxic antiphage defense proteins inhibited by intragenic antitoxin proteins.Proc Natl Acad Sci U S A. 2023 Aug;120(31):e2307382120. doi: 10.1073/pnas.2307382120. Epub 2023 Jul 24. Proc Natl Acad Sci U S A. 2023. PMID: 37487082 Free PMC article.
Abstract
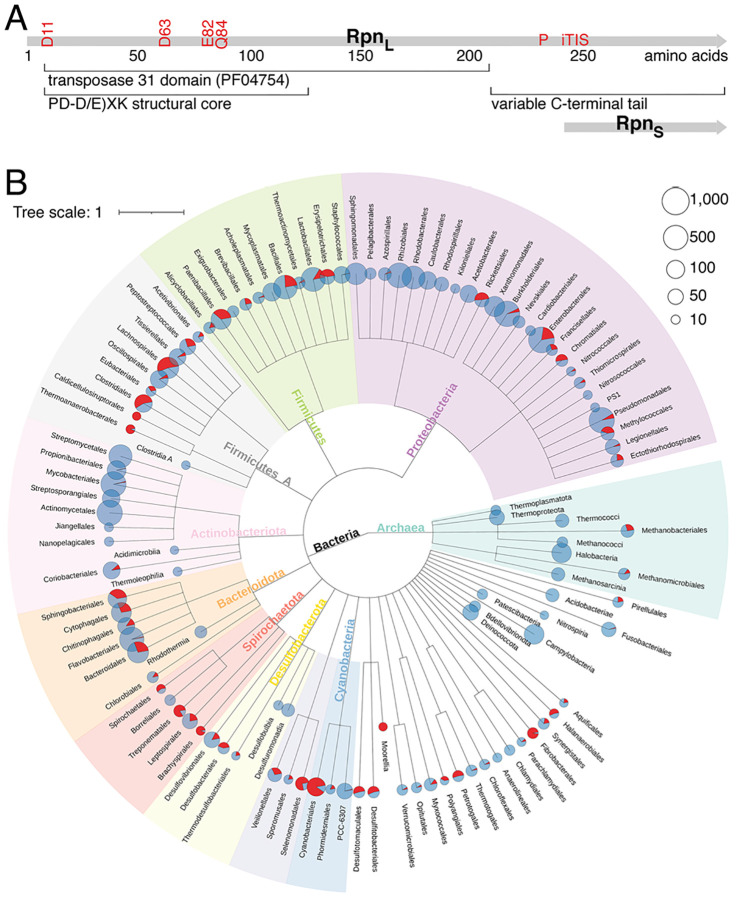
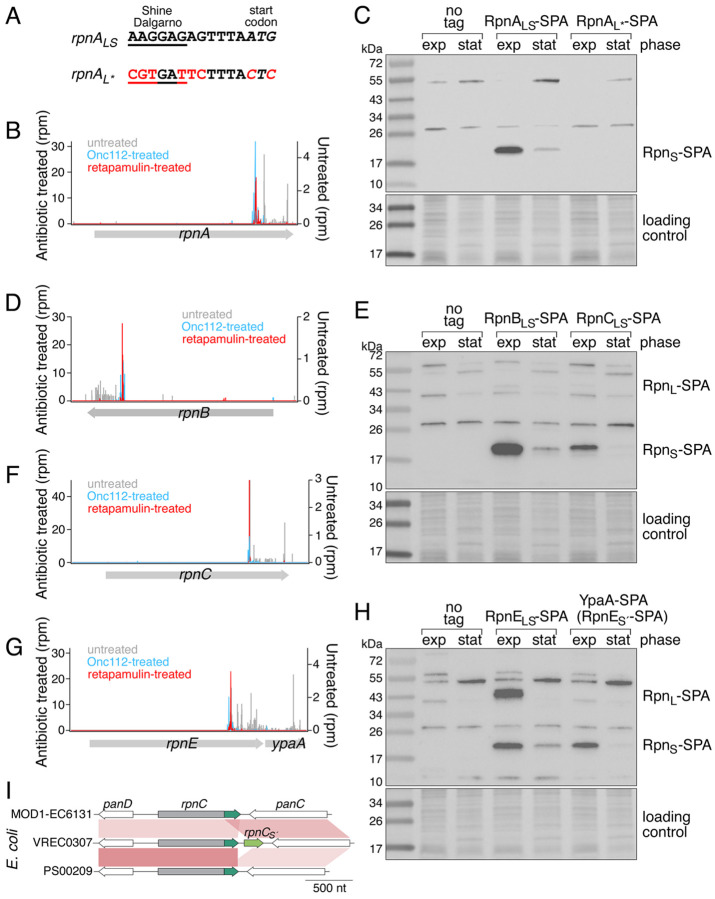
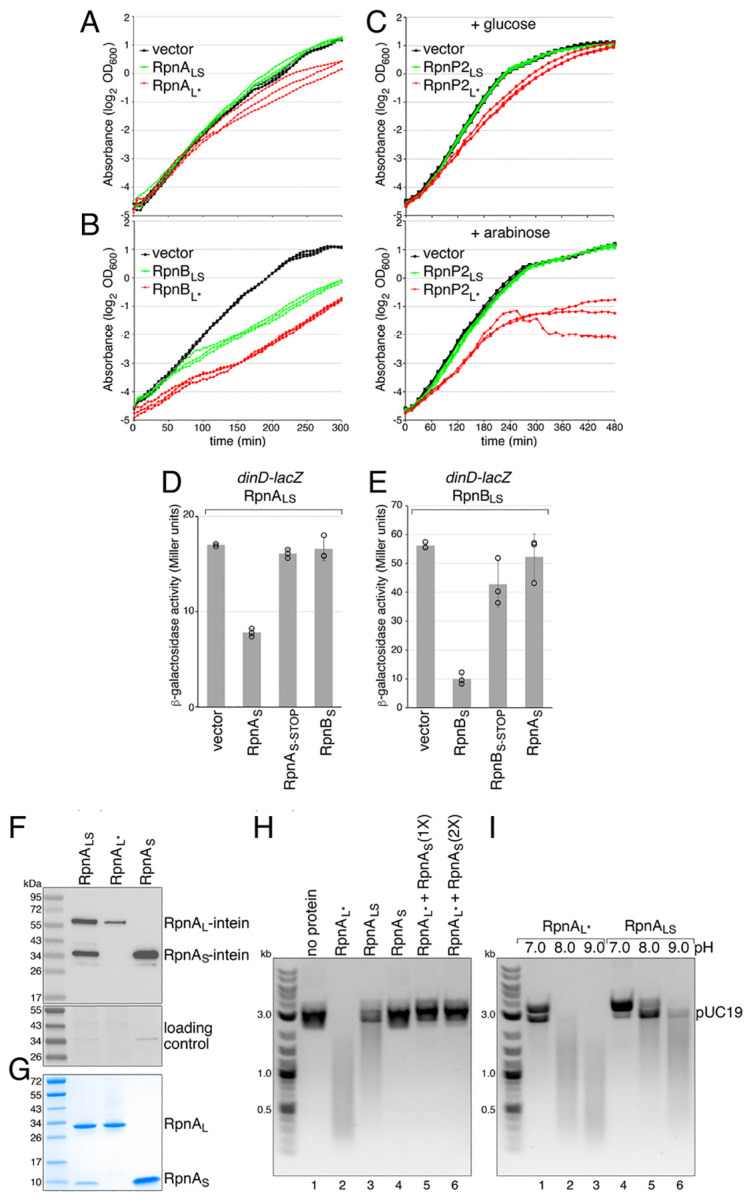
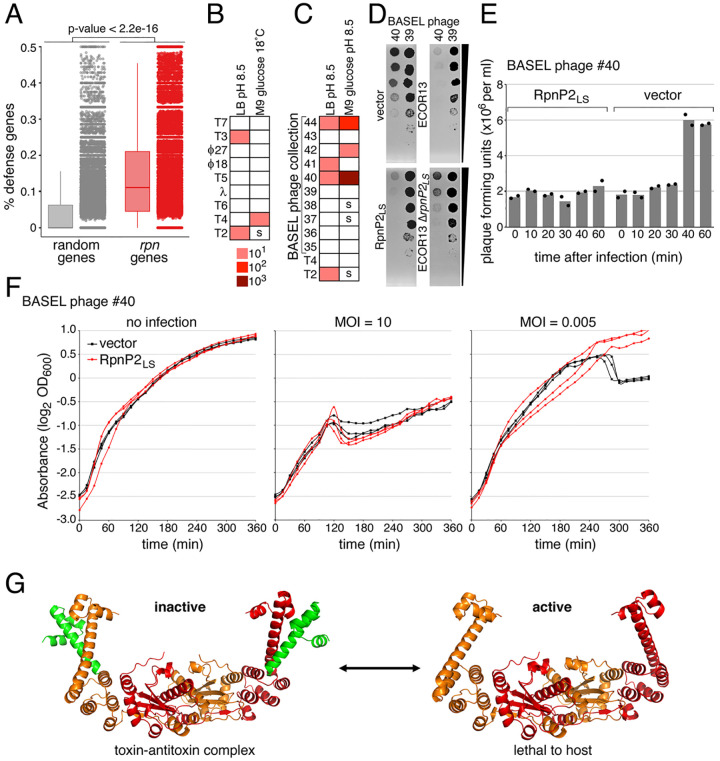
Recombination-promoting nuclease (Rpn) proteins are broadly distributed across bacterial phyla, yet their functions remain unclear. Here we report these proteins are new toxin-antitoxin systems, comprised of genes-within-genes, that combat phage infection. We show the small, highly variable Rpn C -terminal domains (Rpn S ), which are translated separately from the full-length proteins (Rpn L ), directly block the activities of the toxic full-length proteins. The crystal structure of RpnA S revealed a dimerization interface encompassing a helix that can have four amino acid repeats whose number varies widely among strains of the same species. Consistent with strong selection for the variation, we document plasmid-encoded RpnP2 L protects Escherichia coli against certain phages. We propose many more intragenic-encoded proteins that serve regulatory roles remain to be discovered in all organisms.
Significance: Here we document the function of small genes-within-genes, showing they encode antitoxin proteins that block the functions of the toxic DNA endonuclease proteins encoded by the longer rpn genes. Intriguingly, a sequence present in both long and short protein shows extensive variation in the number of four amino acid repeats. Consistent with a strong selection for the variation, we provide evidence that the Rpn proteins represent a phage defense system.
Figures

References
-
- McBride T. M. et al. , Diverse CRISPR-Cas complexes require independent translation of small and large subunits from a single gene. Mol. Cell 80, 971–979 (2020). - PubMed
Publication types
LinkOut - more resources
Full Text Sources