

This is a preprint.
Multi-omics characterization of partial chemical reprogramming reveals evidence of cell rejuvenation
- PMID: 37425825
- PMCID: PMC10327104
- DOI: 10.1101/2023.06.30.546730
Multi-omics characterization of partial chemical reprogramming reveals evidence of cell rejuvenation
Update in
-
Multi-omics characterization of partial chemical reprogramming reveals evidence of cell rejuvenation.Elife. 2024 Mar 22;12:RP90579. doi: 10.7554/eLife.90579. Elife. 2024. PMID: 38517750 Free PMC article.
Abstract
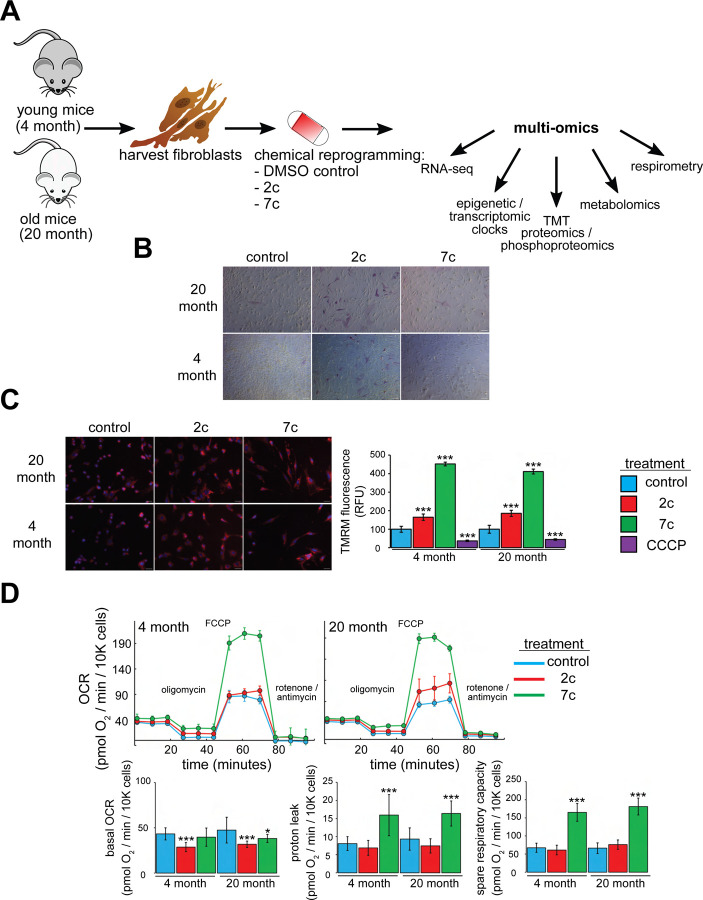
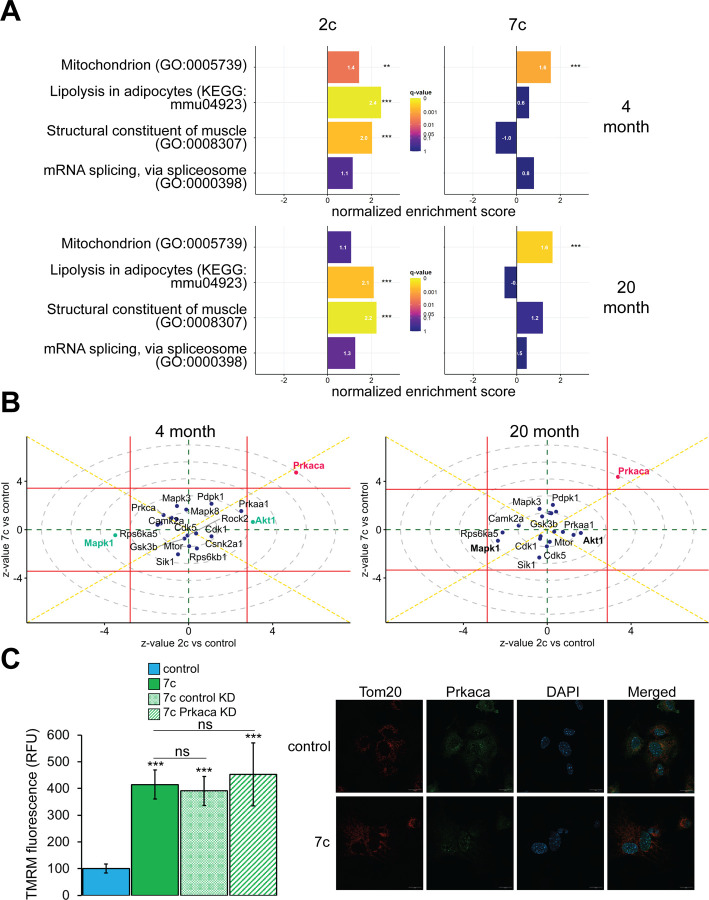
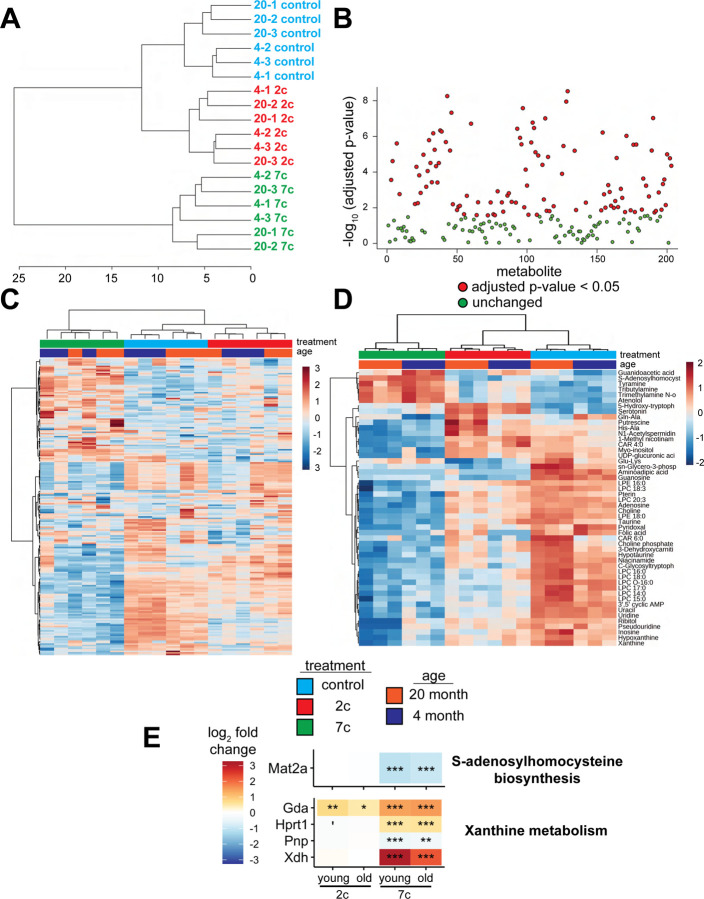
Partial reprogramming by cyclic short-term expression of Yamanaka factors holds promise for shifting cells to younger states and consequently delaying the onset of many diseases of aging. However, the delivery of transgenes and potential risk of teratoma formation present challenges for in vivo applications. Recent advances include the use of cocktails of compounds to reprogram somatic cells, but the characteristics and mechanisms of partial cellular reprogramming by chemicals remain unclear. Here, we report a multi-omics characterization of partial chemical reprogramming in fibroblasts from young and aged mice. We measured the effects of partial chemical reprogramming on the epigenome, transcriptome, proteome, phosphoproteome, and metabolome. At the transcriptome, proteome, and phosphoproteome levels, we saw widescale changes induced by this treatment, with the most notable signature being an upregulation of mitochondrial oxidative phosphorylation. Furthermore, at the metabolome level, we observed a reduction in the accumulation of aging-related metabolites. Using both transcriptomic and epigenetic clock-based analyses, we show that partial chemical reprogramming reduces the biological age of mouse fibroblasts. We demonstrate that these changes have functional impacts, as evidenced by changes in cellular respiration and mitochondrial membrane potential. Taken together, these results illuminate the potential for chemical reprogramming reagents to rejuvenate aged biological systems and warrant further investigation into adapting these approaches for in vivo age reversal.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials