

This is a preprint.
Combinatorial transcription factor binding encodes cis-regulatory wiring of forebrain GABAergic neurogenesis
- PMID: 37425940
- PMCID: PMC10327028
- DOI: 10.1101/2023.06.28.546894
Combinatorial transcription factor binding encodes cis-regulatory wiring of forebrain GABAergic neurogenesis
Update in
-
Combinatorial transcription factor binding encodes cis-regulatory wiring of mouse forebrain GABAergic neurogenesis.Dev Cell. 2025 Jan 20;60(2):288-304.e6. doi: 10.1016/j.devcel.2024.10.004. Epub 2024 Oct 30. Dev Cell. 2025. PMID: 39481376
Abstract
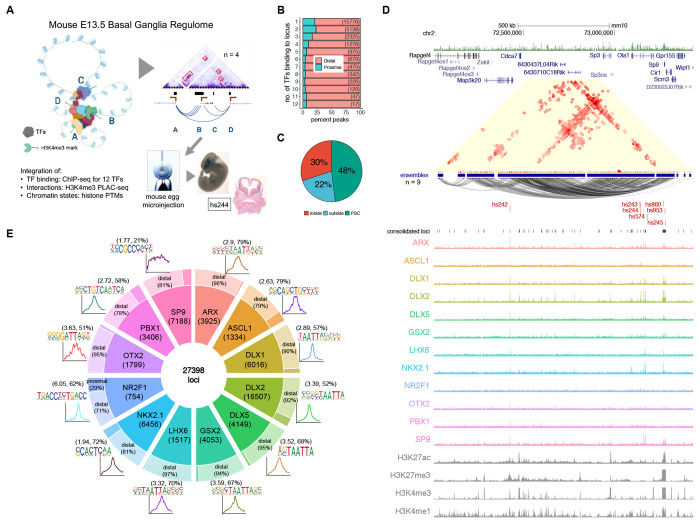
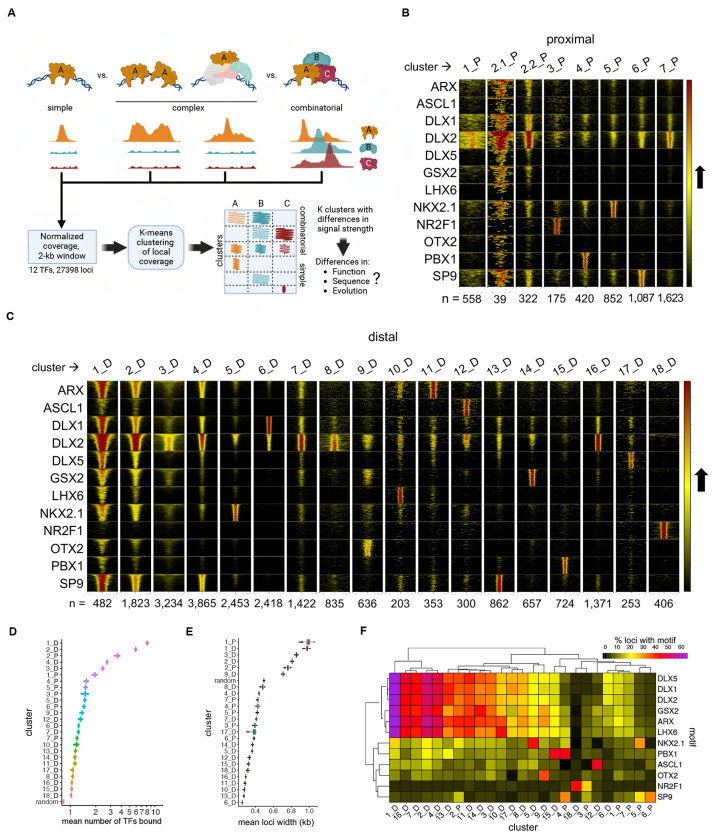
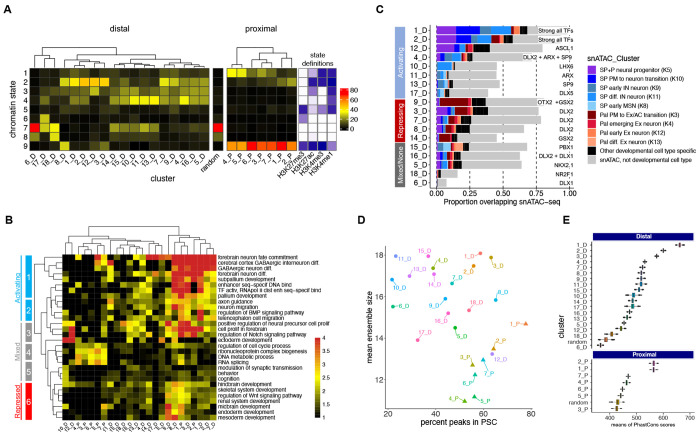
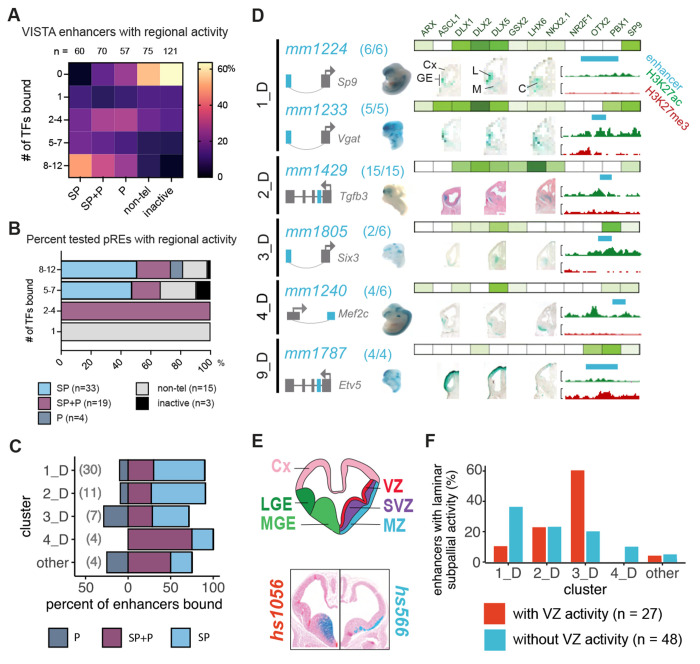
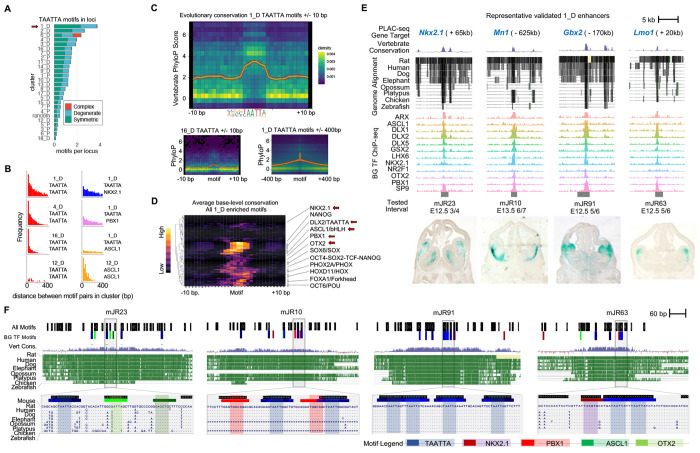
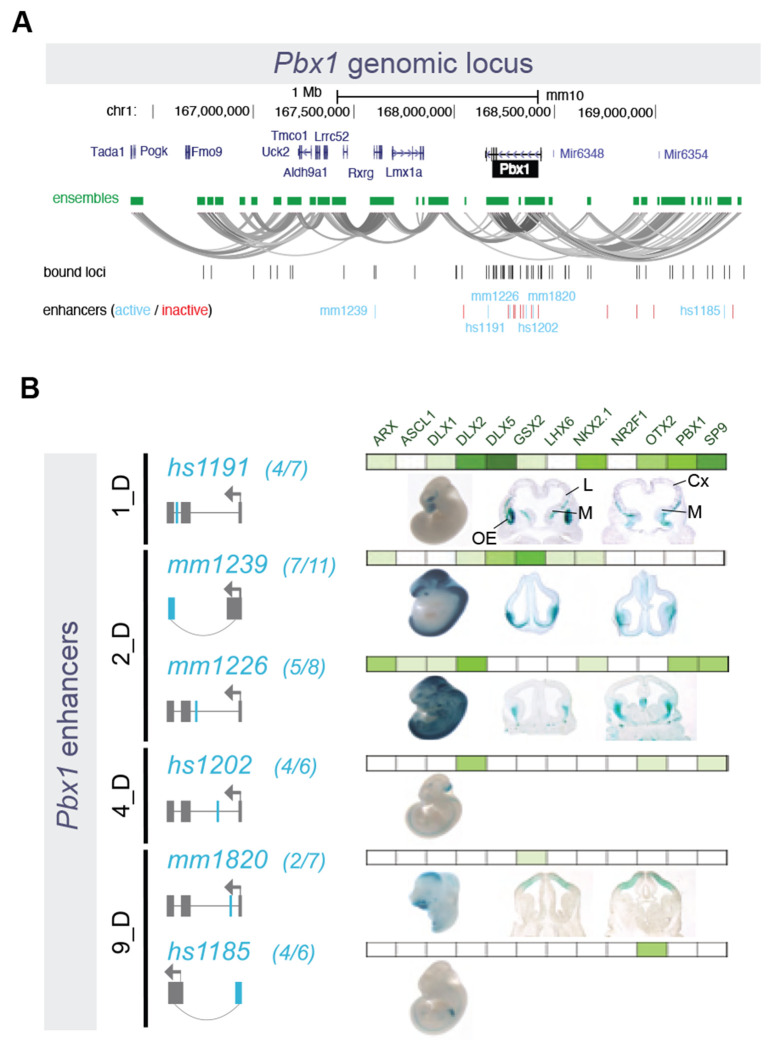
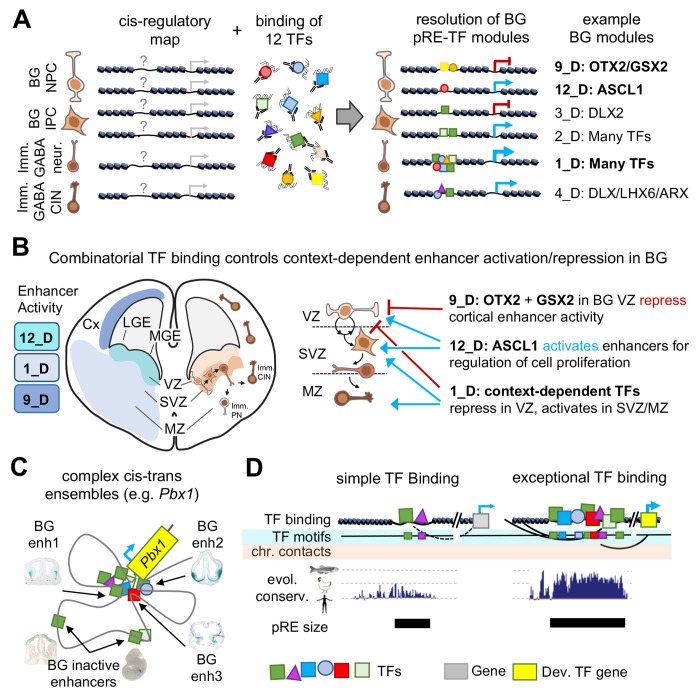
Transcription factors (TFs) bind combinatorially to genomic cis-regulatory elements (cREs), orchestrating transcription programs. While studies of chromatin state and chromosomal interactions have revealed dynamic neurodevelopmental cRE landscapes, parallel understanding of the underlying TF binding lags. To elucidate the combinatorial TF-cRE interactions driving mouse basal ganglia development, we integrated ChIP-seq for twelve TFs, H3K4me3-associated enhancer-promoter interactions, chromatin and transcriptional state, and transgenic enhancer assays. We identified TF-cREs modules with distinct chromatin features and enhancer activity that have complementary roles driving GABAergic neurogenesis and suppressing other developmental fates. While the majority of distal cREs were bound by one or two TFs, a small proportion were extensively bound, and these enhancers also exhibited exceptional evolutionary conservation, motif density, and complex chromosomal interactions. Our results provide new insights into how modules of combinatorial TF-cRE interactions activate and repress developmental expression programs and demonstrate the value of TF binding data in modeling gene regulatory wiring.
Conflict of interest statement
STATEMENT OF COMPETING INTERESTS J.L.R.R. is co-founder, stockholder, and currently on the scientific board of Neurona, a company studying the potential therapeutic use of interneuron transplantation. The other authors declare no competing interests.
Figures
References
-
- Rubenstein J. L. R.& Campbell K. Chapter 18 - Neurogenesis in the basal ganglia. in (eds. Rubenstein J., Rakic P., Chen B. & Kwan K. Y. B. T.-P. and S C. T. in the S. D. C. N. and S. P. N.. (Second E.) 399–426 (Academic Press, 2020). doi:10.1016/B978-0-12-814405-3.00018-7. - DOI
-
- Grillner S. & Robertson B. The Basal Ganglia Over 500 Million Years. Curr. Biol. 26, R1088–R1100 (2016). - PubMed
-
- Stephenson-Jones M., Samuelsson E., Ericsson J., Robertson B. & Grillner S. Evolutionary Conservation of the Basal Ganglia as a Common Vertebrate Mechanism for Action Selection. Curr. Biol. 21, 1081–1091 (2011). - PubMed
-
- Holland P. W. H. & Takahashi T. The evolution of homeobox genes: Implications for the study of brain development. Brain Res. Bull. 66, 484–490 (2005). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous