

This is a preprint.
Cell Type- and Tissue-specific Enhancers in Craniofacial Development
- PMID: 37425964
- PMCID: PMC10327103
- DOI: 10.1101/2023.06.26.546603
Cell Type- and Tissue-specific Enhancers in Craniofacial Development
Abstract
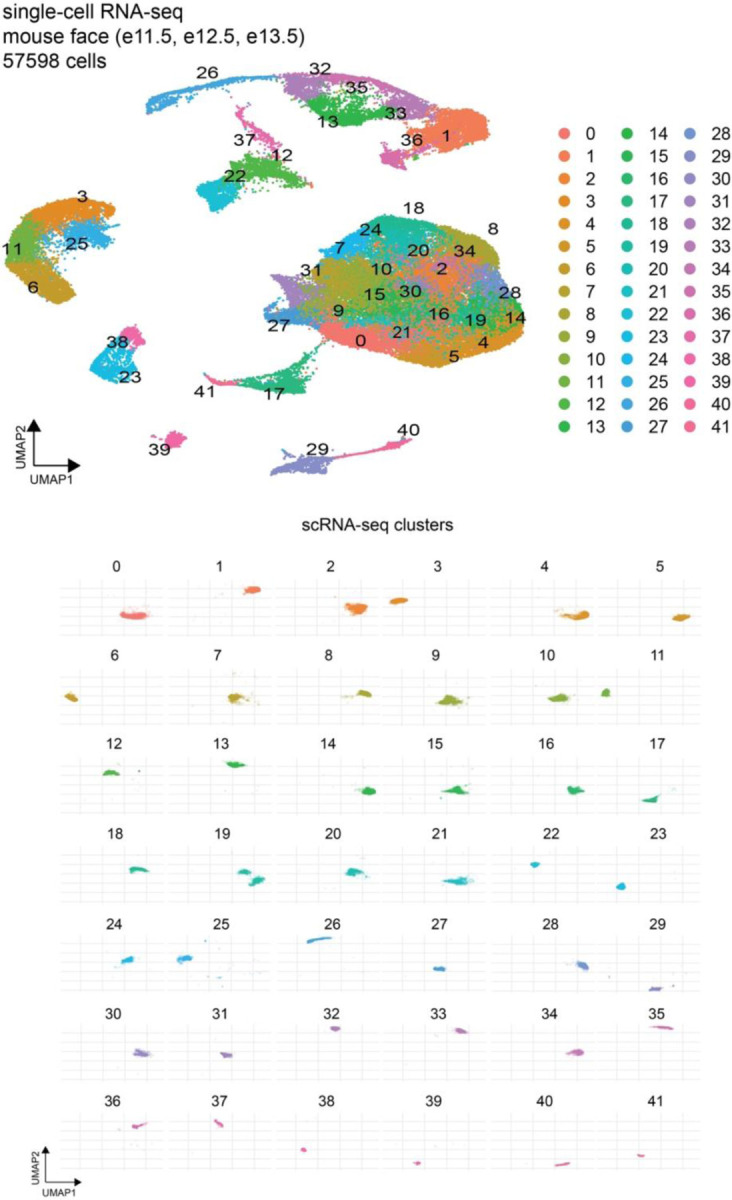
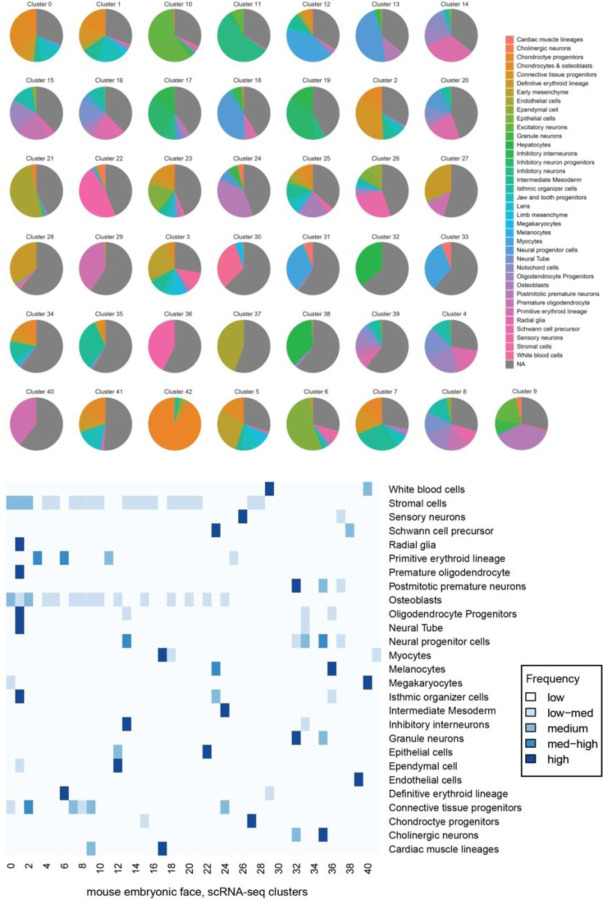
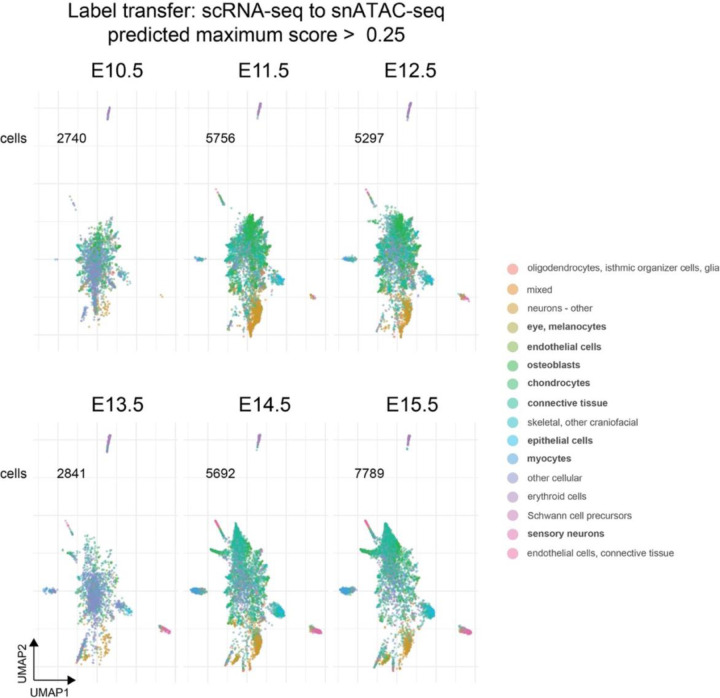
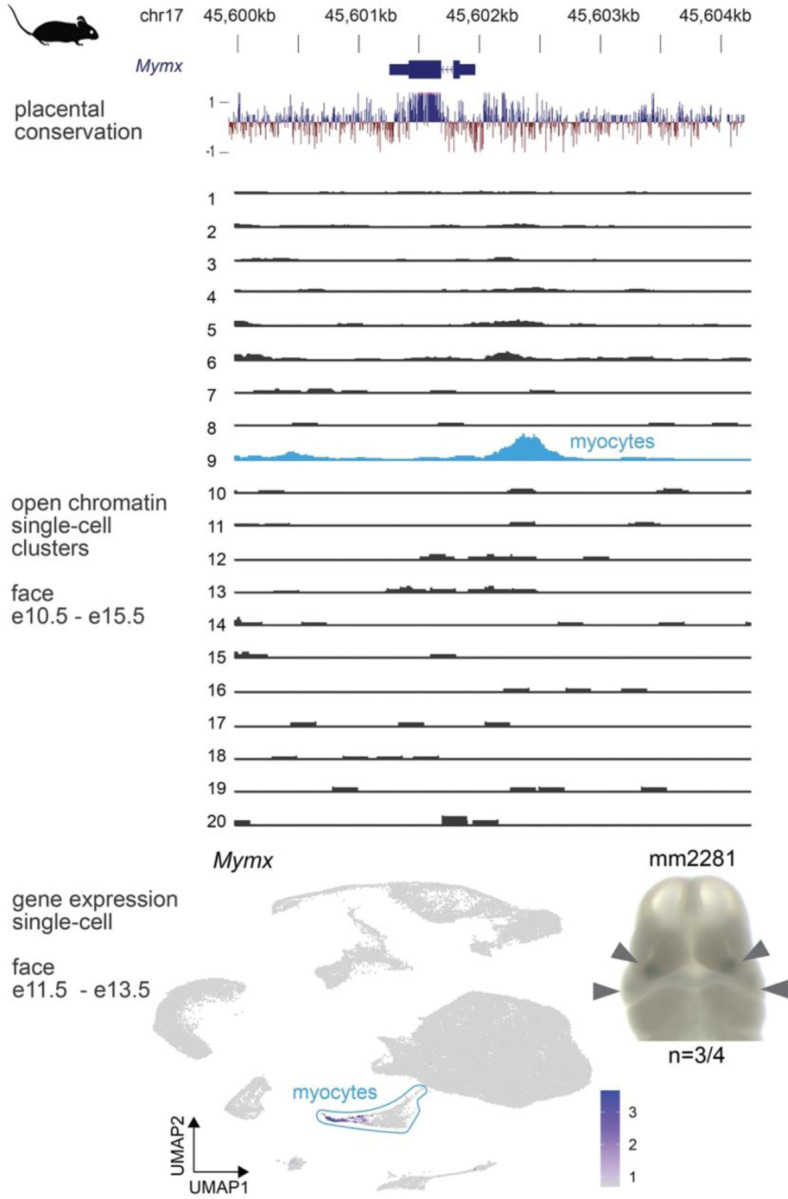
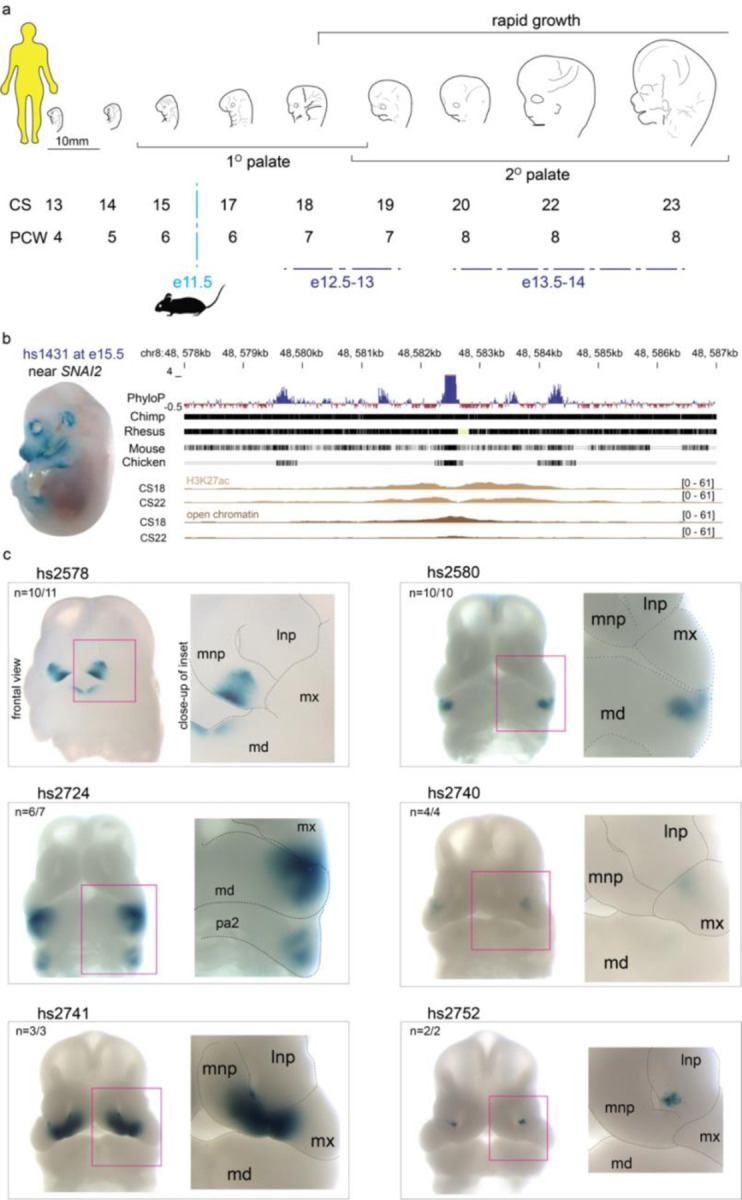
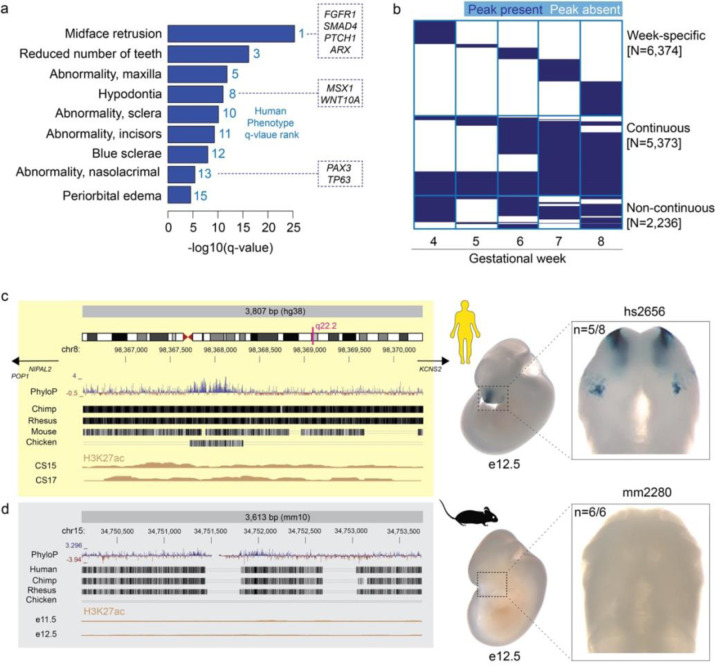
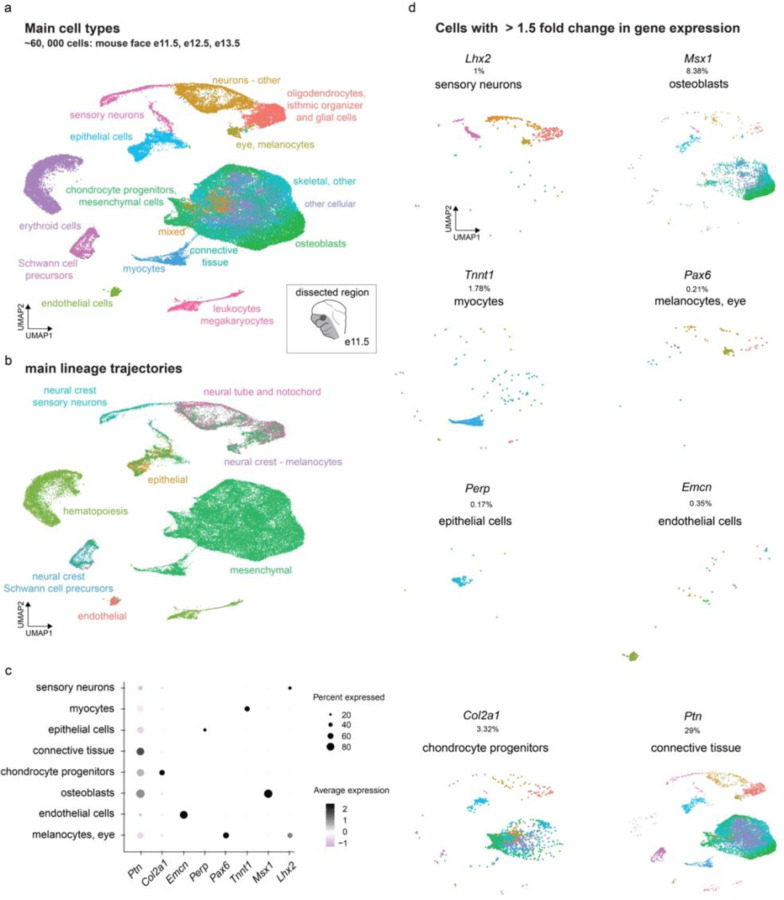
The genetic basis of craniofacial birth defects and general variation in human facial shape remains poorly understood. Distant-acting transcriptional enhancers are a major category of non-coding genome function and have been shown to control the fine-tuned spatiotemporal expression of genes during critical stages of craniofacial development1-3. However, a lack of accurate maps of the genomic location and cell type-specific in vivo activities of all craniofacial enhancers prevents their systematic exploration in human genetics studies. Here, we combined histone modification and chromatin accessibility profiling from different stages of human craniofacial development with single-cell analyses of the developing mouse face to create a comprehensive catalogue of the regulatory landscape of facial development at tissue- and single cell-resolution. In total, we identified approximately 14,000 enhancers across seven developmental stages from weeks 4 through 8 of human embryonic face development. We used transgenic mouse reporter assays to determine the in vivo activity patterns of human face enhancers predicted from these data. Across 16 in vivo validated human enhancers, we observed a rich diversity of craniofacial subregions in which these enhancers are active in vivo. To annotate the cell type specificities of human-mouse conserved enhancers, we performed single-cell RNA-seq and single-nucleus ATAC-seq of mouse craniofacial tissues from embryonic days e11.5 to e15.5. By integrating these data across species, we find that the majority (56%) of human craniofacial enhancers are functionally conserved in mice, providing cell type- and embryonic stage-resolved predictions of their in vivo activity profiles. Using retrospective analysis of known craniofacial enhancers in combination with single cell-resolved transgenic reporter assays, we demonstrate the utility of these data for predicting the in vivo cell type specificity of enhancers. Taken together, our data provide an expansive resource for genetic and developmental studies of human craniofacial development.
Conflict of interest statement
Declaration of Interests Bing Ren is a co-founder of Arima Genomics, Inc, and Epigenome Technologies, Inc.
Figures










References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials