

Lipid droplets are versatile organelles involved in plant development and plant response to environmental changes
- PMID: 37426978
- PMCID: PMC10327486
- DOI: 10.3389/fpls.2023.1193905
Lipid droplets are versatile organelles involved in plant development and plant response to environmental changes
Abstract
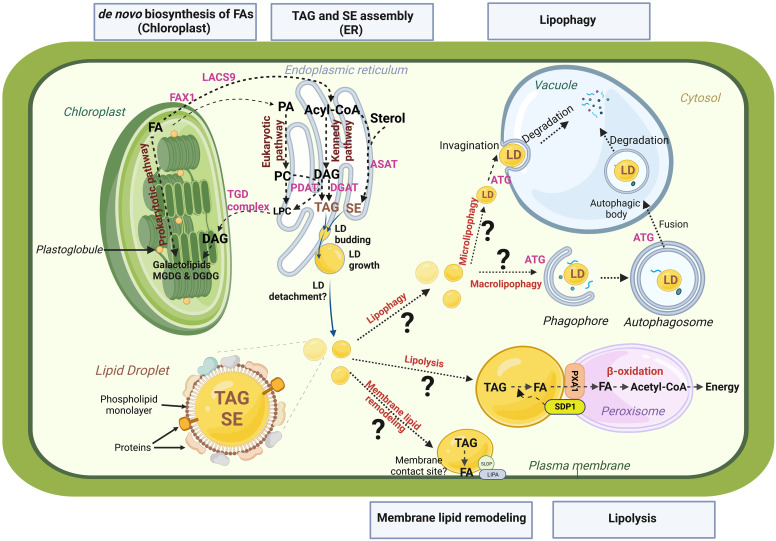
Since decades plant lipid droplets (LDs) are described as storage organelles accumulated in seeds to provide energy for seedling growth after germination. Indeed, LDs are the site of accumulation for neutral lipids, predominantly triacylglycerols (TAGs), one of the most energy-dense molecules, and sterol esters. Such organelles are present in the whole plant kingdom, from microalgae to perennial trees, and can probably be found in all plant tissues. Several studies over the past decade have revealed that LDs are not merely simple energy storage compartments, but also dynamic structures involved in diverse cellular processes like membrane remodeling, regulation of energy homeostasis and stress responses. In this review, we aim to highlight the functions of LDs in plant development and response to environmental changes. In particular, we tackle the fate and roles of LDs during the plant post-stress recovery phase.
Keywords: autophagy; environmental stress; heat; lipid droplets; lipolysis; membrane remodeling; post-stress recovery; triacylglycerol.
Copyright © 2023 Bouchnak, Coulon, Salis, D’Andréa and Bréhélin.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Biogenesis and Lipase-Mediated Mobilization of Lipid Droplets in Plants.Plants (Basel). 2022 May 5;11(9):1243. doi: 10.3390/plants11091243. Plants (Basel). 2022. PMID: 35567244 Free PMC article. Review.
-
Lipid droplets degradation mechanisms from microalgae to mammals, a comparative overview.Biochimie. 2024 Dec;227(Pt B):19-34. doi: 10.1016/j.biochi.2024.09.006. Epub 2024 Sep 17. Biochimie. 2024. PMID: 39299537 Review.
-
Degradation of Lipid Droplets in Plants and Algae-Right Time, Many Paths, One Goal.Front Plant Sci. 2020 Sep 9;11:579019. doi: 10.3389/fpls.2020.579019. eCollection 2020. Front Plant Sci. 2020. PMID: 33014002 Free PMC article. Review.
-
Lipid droplet dynamics in budding yeast.Cell Mol Life Sci. 2015 Jul;72(14):2677-95. doi: 10.1007/s00018-015-1903-5. Epub 2015 Apr 18. Cell Mol Life Sci. 2015. PMID: 25894691 Free PMC article. Review.
-
Here, there and everywhere - the importance of neutral lipids in plant growth and development.Postepy Biochem. 2021 Nov 12;68(1):46-56. doi: 10.18388/pb.2021_409. Print 2022 Mar 31. Postepy Biochem. 2021. PMID: 35569044 Review.
Cited by
-
Lipid droplets: New roles as mediators of biotic and abiotic stress.Plant Physiol. 2024 Dec 23;197(1):kiae340. doi: 10.1093/plphys/kiae340. Plant Physiol. 2024. PMID: 38888997 Free PMC article. No abstract available.
-
Molecular characterization of CeOLE6, a diverged SH oleosin gene, preferentially expressed in Cyperus esculentus tubers.Planta. 2024 Oct 23;260(6):122. doi: 10.1007/s00425-024-04553-5. Planta. 2024. PMID: 39438351
-
Inducible expression of DEFECTIVE IN ANTHER DEHISCENCE 1 enhances triacylglycerol accumulation and lipid droplet formation in vegetative tissues.Plant J. 2025 Mar;121(5):e70088. doi: 10.1111/tpj.70088. Plant J. 2025. PMID: 40052427 Free PMC article.
-
Degeneration of oil bodies by rough endoplasmic reticulum -associated protein during seed germination in Cannabis sativa.AoB Plants. 2023 Nov 22;15(6):plad082. doi: 10.1093/aobpla/plad082. eCollection 2023 Dec. AoB Plants. 2023. PMID: 38094511 Free PMC article.
-
The jojoba lipid droplet protein LDAP1 facilitates the packaging of wax esters into lipid droplets.Plant Cell. 2025 Aug 1;37(8):koaf115. doi: 10.1093/plcell/koaf115. Plant Cell. 2025. PMID: 40794685 Free PMC article.
References
-
- Avin-Wittenberg T., Bajdzienko K., Wittenberg G., Alseekh S., Tohge T., Bock R., et al. . (2015). Global analysis of the role of autophagy in cellular metabolism and energy homeostasis in arabidopsis seedlings under carbon starvation. Plant Cell 27, 306–322. doi: 10.1105/tpc.114.134205 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous