

Multiomics of Tissue Extracellular Vesicles Identifies Unique Modulators of Atherosclerosis and Calcific Aortic Valve Stenosis
- PMID: 37427430
- PMCID: PMC10527599
- DOI: 10.1161/CIRCULATIONAHA.122.063402
Multiomics of Tissue Extracellular Vesicles Identifies Unique Modulators of Atherosclerosis and Calcific Aortic Valve Stenosis
Abstract
Background: Fewer than 50% of patients who develop aortic valve calcification have concomitant atherosclerosis, implying differential pathogenesis. Although circulating extracellular vesicles (EVs) act as biomarkers of cardiovascular diseases, tissue-entrapped EVs are associated with early mineralization, but their cargoes, functions, and contributions to disease remain unknown.
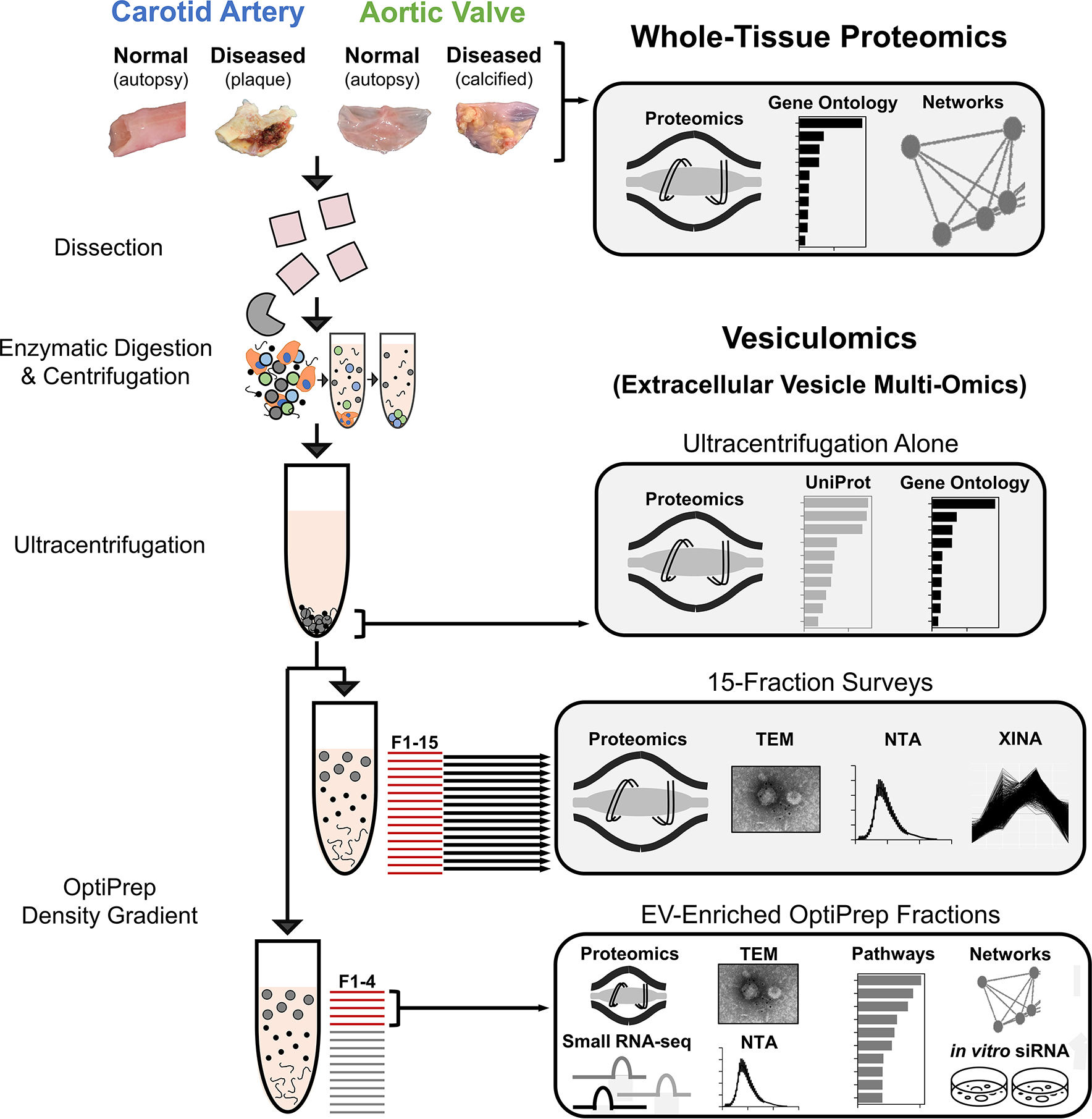
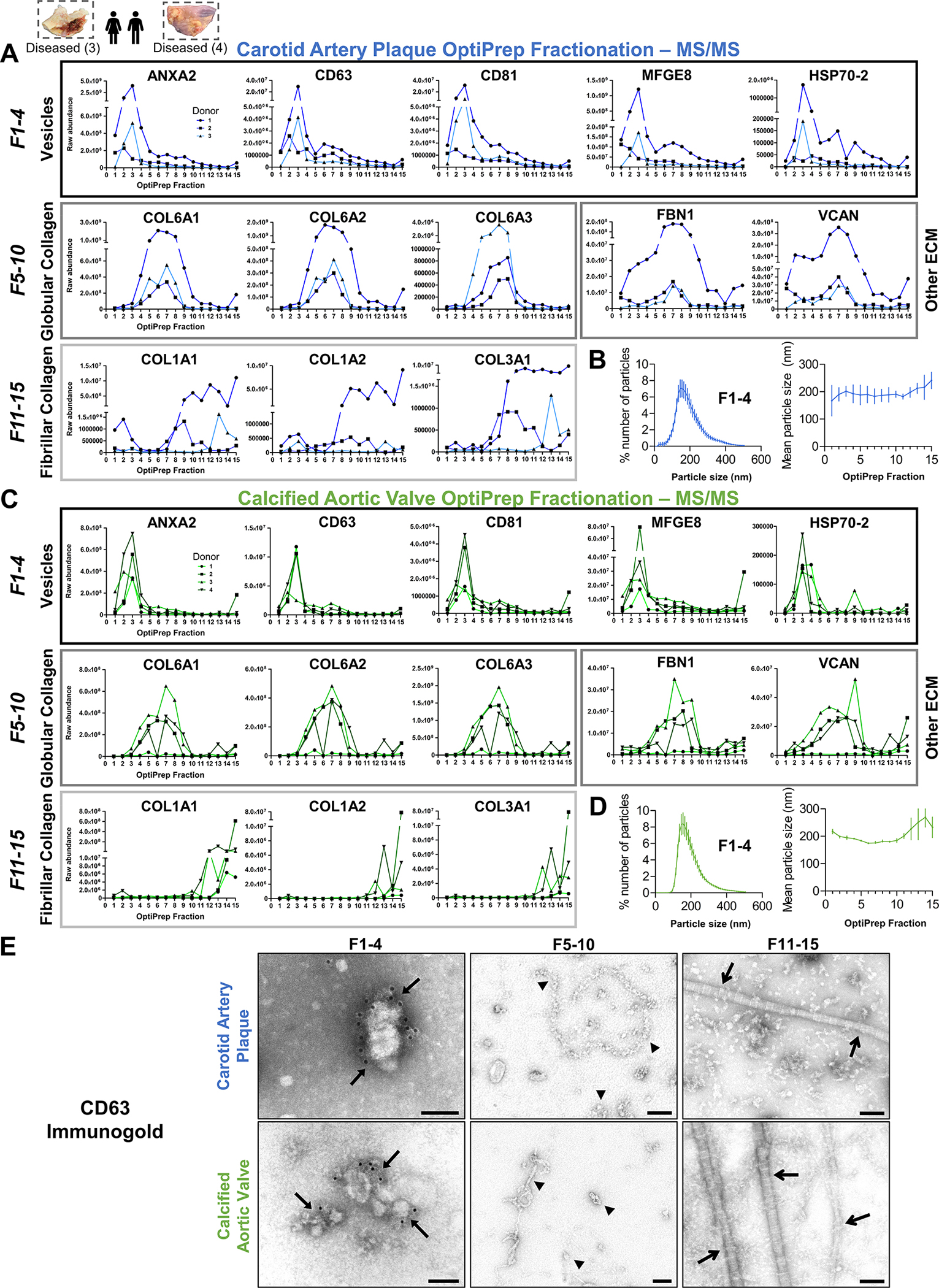
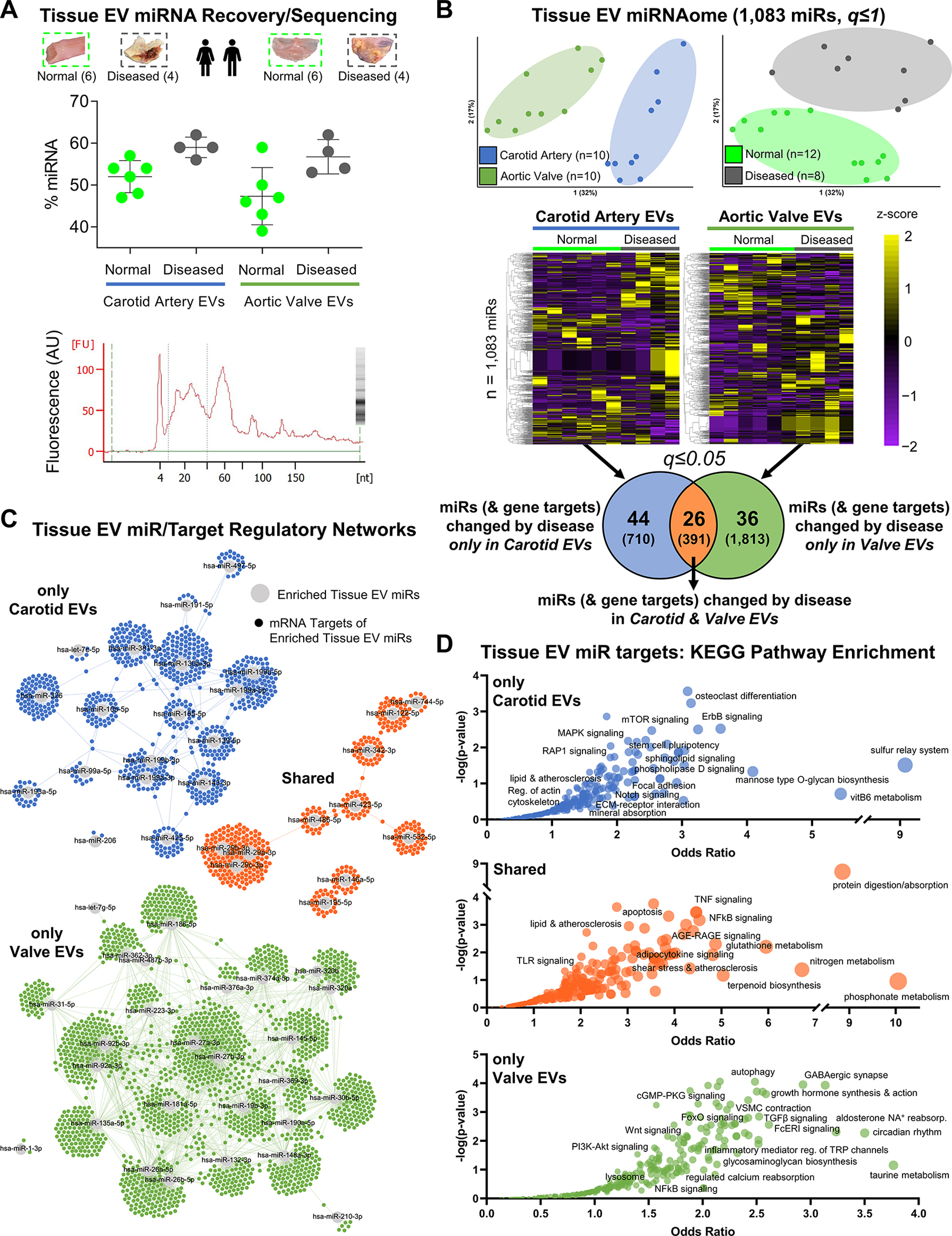
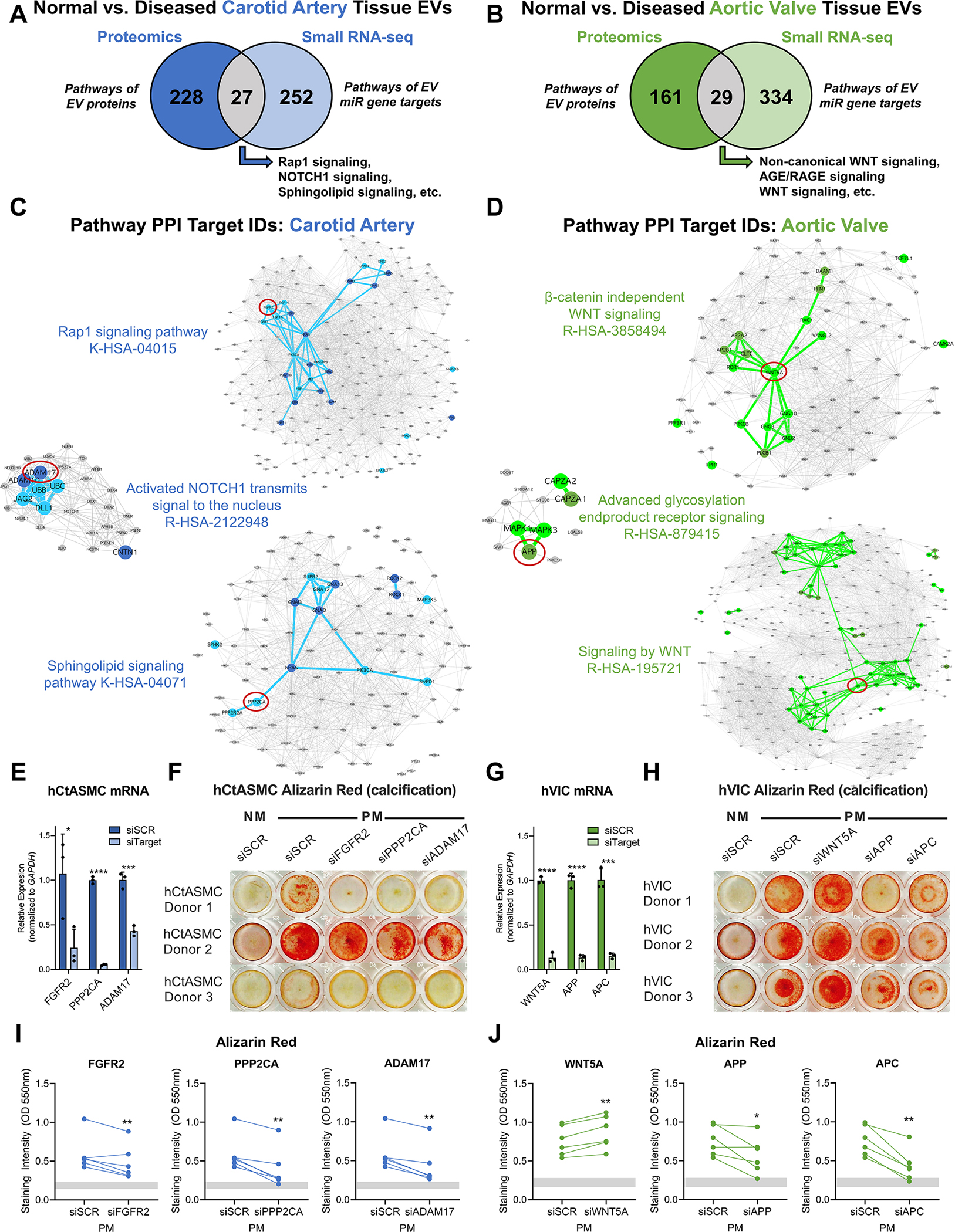
Methods: Disease stage-specific proteomics was performed on human carotid endarterectomy specimens (n=16) and stenotic aortic valves (n=18). Tissue EVs were isolated from human carotid arteries (normal, n=6; diseased, n=4) and aortic valves (normal, n=6; diseased, n=4) by enzymatic digestion, (ultra)centrifugation, and a 15-fraction density gradient validated by proteomics, CD63-immunogold electron microscopy, and nanoparticle tracking analysis. Vesiculomics, comprising vesicular proteomics and small RNA-sequencing, was conducted on tissue EVs. TargetScan identified microRNA targets. Pathway network analyses prioritized genes for validation in primary human carotid artery smooth muscle cells and aortic valvular interstitial cells.
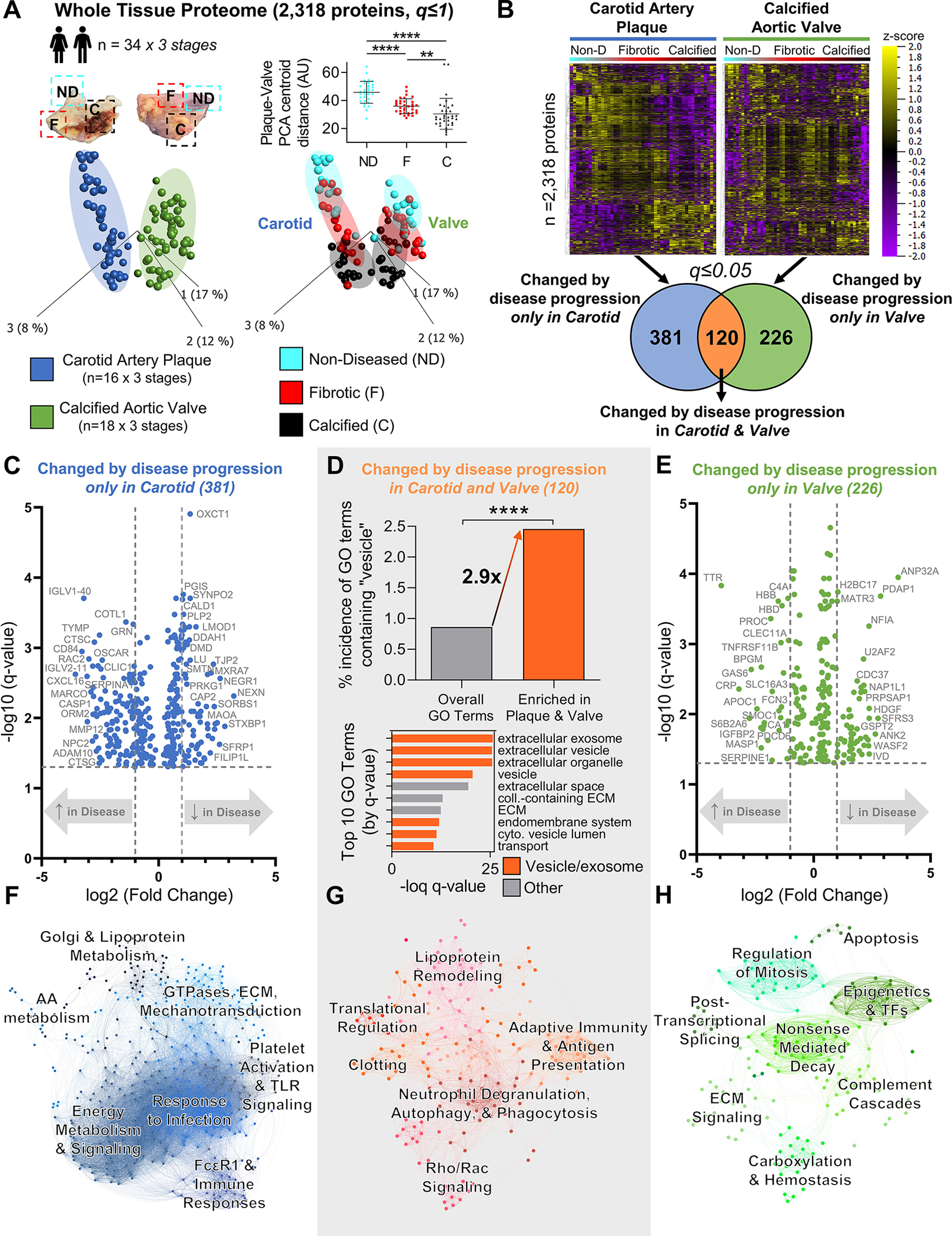
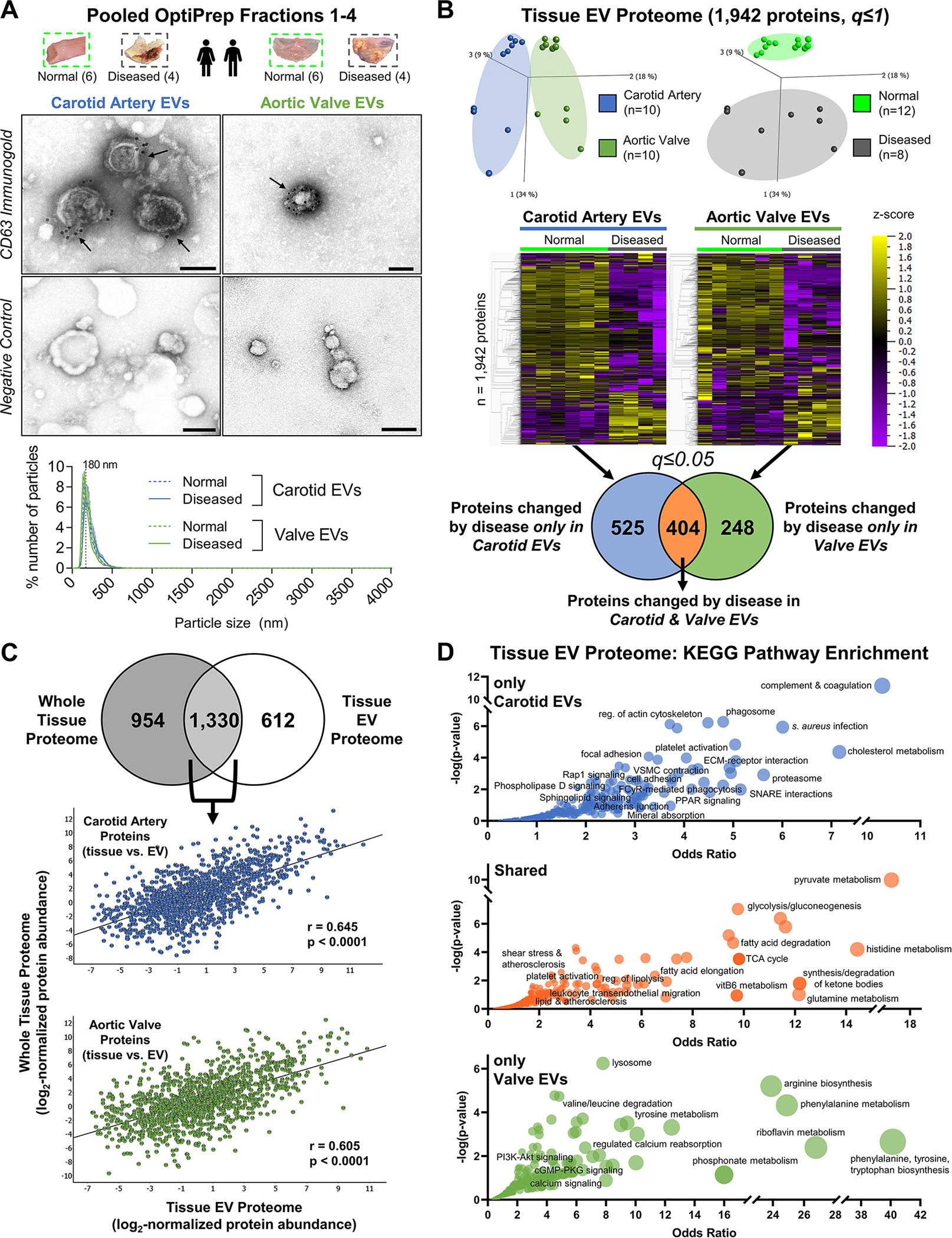
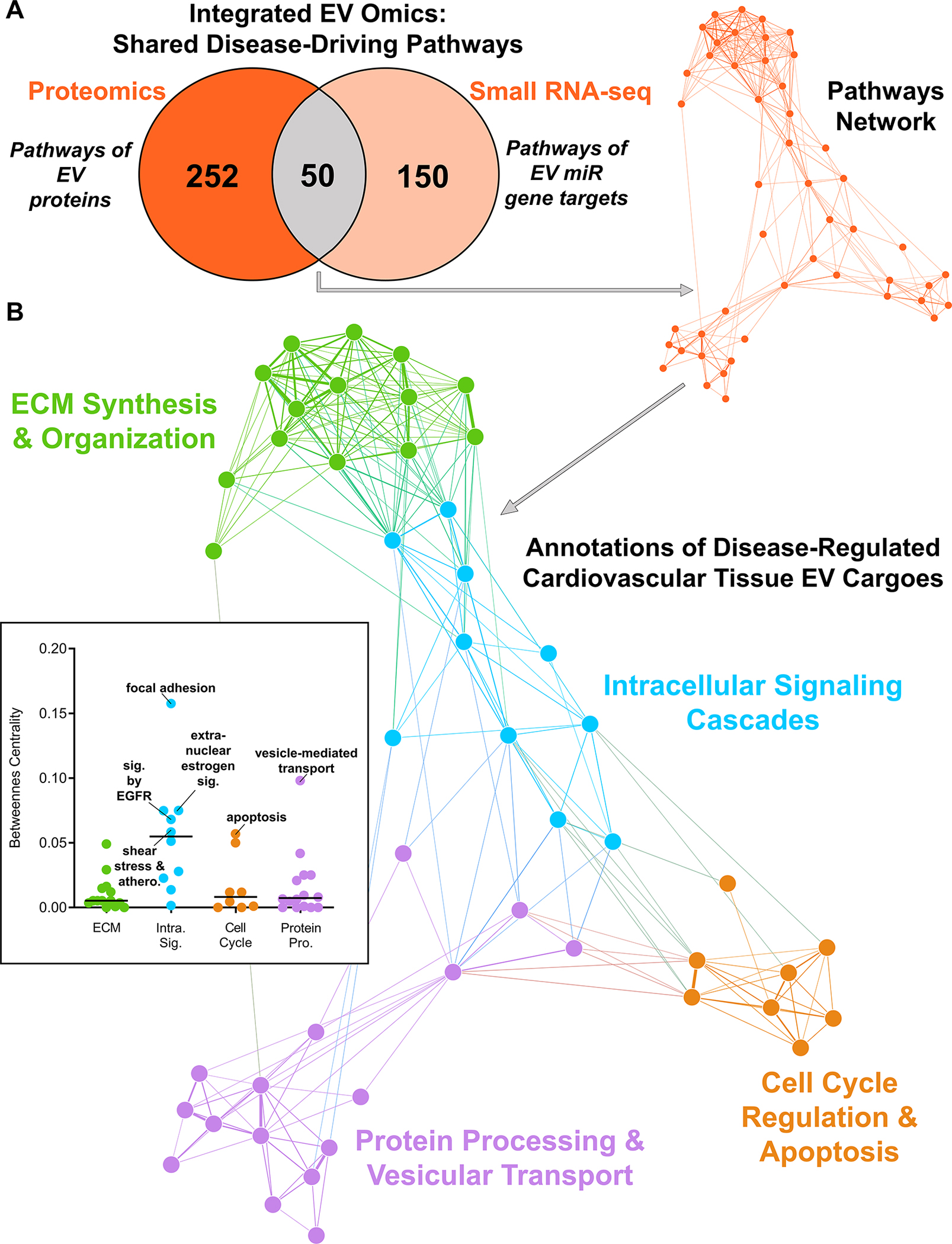
Results: Disease progression drove significant convergence (P<0.0001) of carotid artery plaque and calcified aortic valve proteomes (2318 proteins). Each tissue also retained a unique subset of differentially enriched proteins (381 in plaques; 226 in valves; q<0.05). Vesicular gene ontology terms increased 2.9-fold (P<0.0001) among proteins modulated by disease in both tissues. Proteomics identified 22 EV markers in tissue digest fractions. Networks of proteins and microRNA targets changed by disease progression in both artery and valve EVs revealed shared involvement in intracellular signaling and cell cycle regulation. Vesiculomics identified 773 proteins and 80 microRNAs differentially enriched by disease exclusively in artery or valve EVs (q<0.05); multiomics integration found tissue-specific EV cargoes associated with procalcific Notch and Wnt signaling in carotid arteries and aortic valves, respectively. Knockdown of tissue-specific EV-derived molecules FGFR2, PPP2CA, and ADAM17 in human carotid artery smooth muscle cells and WNT5A, APP, and APC in human aortic valvular interstitial cells significantly modulated calcification.
Conclusions: The first comparative proteomics study of human carotid artery plaques and calcified aortic valves identifies unique drivers of atherosclerosis versus aortic valve stenosis and implicates EVs in advanced cardiovascular calcification. We delineate a vesiculomics strategy to isolate, purify, and study protein and RNA cargoes from EVs entrapped in fibrocalcific tissues. Integration of vesicular proteomics and transcriptomics by network approaches revealed novel roles for tissue EVs in modulating cardiovascular disease.
Keywords: aortic valve stenosis; atherosclerosis; extracellular vesicles; microRNAs; proteomics.
Conflict of interest statement
Figures
References
-
- Chen J, Budoff MJ, Reilly MP, Yang W, Rosas SE, Rahman M, Zhang X, Roy JA, Lustigova E, Nessel L, et al. Coronary Artery Calcification and Risk of Cardiovascular Disease and Death Among Patients With Chronic Kidney Disease. JAMA cardiology. 2017;2:635–643. doi: 10.1001/jamacardio.2017.0363 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
