

Functional interrogation of lymphocyte subsets in alopecia areata using single-cell RNA sequencing
- PMID: 37428932
- PMCID: PMC10629527
- DOI: 10.1073/pnas.2305764120
Functional interrogation of lymphocyte subsets in alopecia areata using single-cell RNA sequencing
Abstract
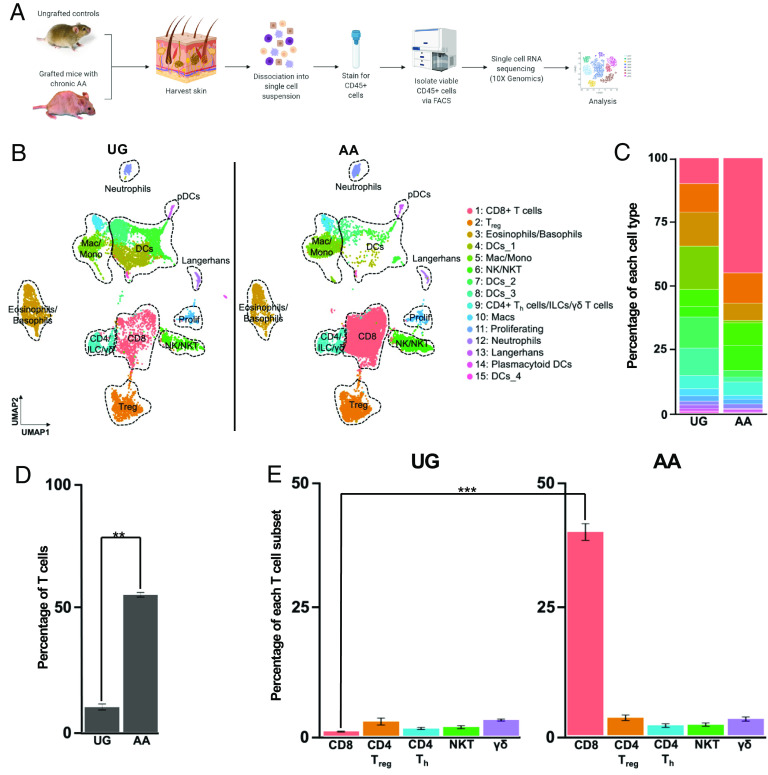
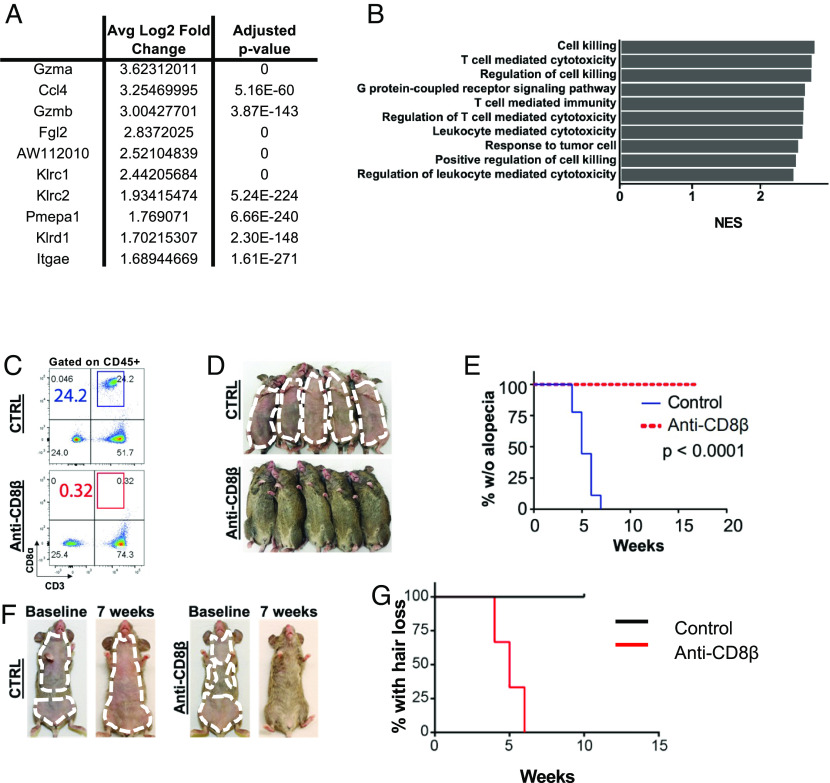
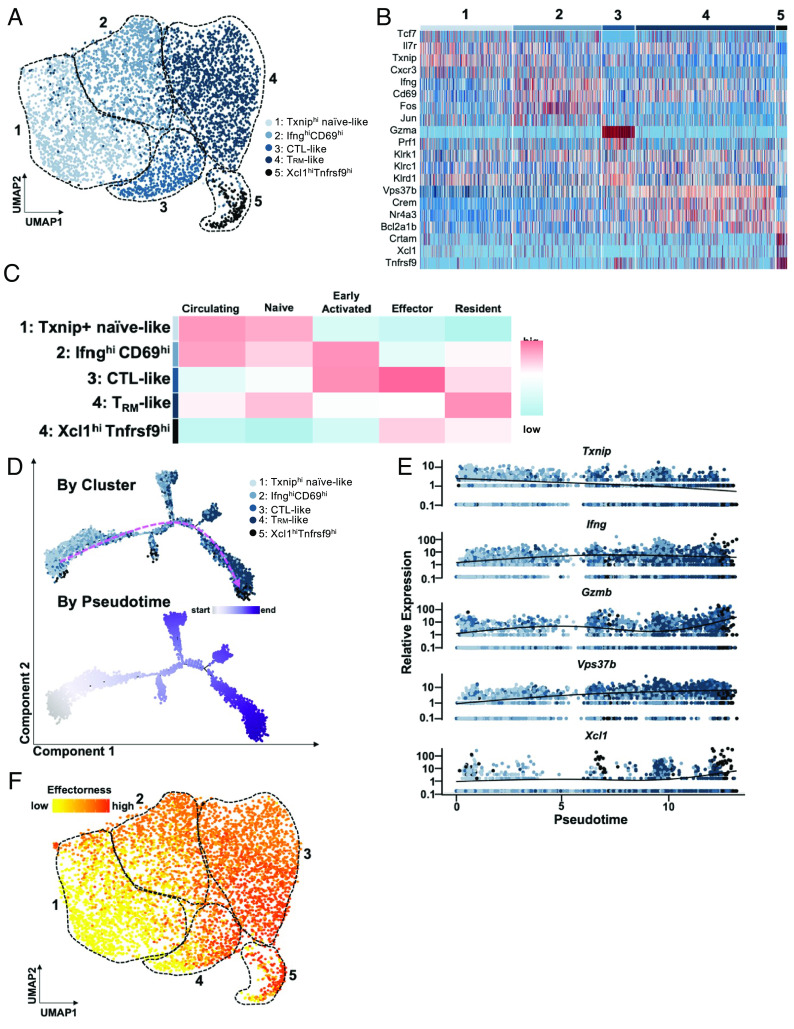
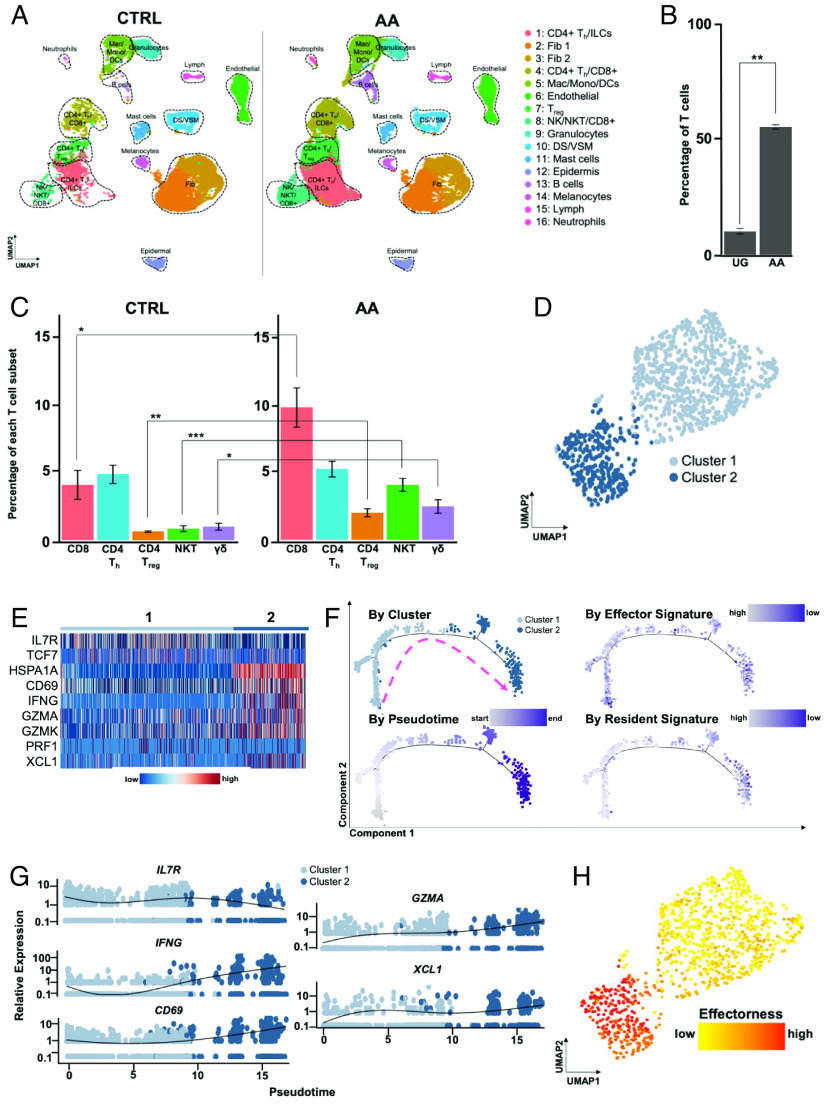
Alopecia areata (AA) is among the most prevalent autoimmune diseases, but the development of innovative therapeutic strategies has lagged due to an incomplete understanding of the immunological underpinnings of disease. Here, we performed single-cell RNA sequencing (scRNAseq) of skin-infiltrating immune cells from the graft-induced C3H/HeJ mouse model of AA, coupled with antibody-based depletion to interrogate the functional role of specific cell types in AA in vivo. Since AA is predominantly T cell-mediated, we focused on dissecting lymphocyte function in AA. Both our scRNAseq and functional studies established CD8+ T cells as the primary disease-driving cell type in AA. Only the depletion of CD8+ T cells, but not CD4+ T cells, NK, B, or γδ T cells, was sufficient to prevent and reverse AA. Selective depletion of regulatory T cells (Treg) showed that Treg are protective against AA in C3H/HeJ mice, suggesting that failure of Treg-mediated immunosuppression is not a major disease mechanism in AA. Focused analyses of CD8+ T cells revealed five subsets, whose heterogeneity is defined by an "effectorness gradient" of interrelated transcriptional states that culminate in increased effector function and tissue residency. scRNAseq of human AA skin showed that CD8+ T cells in human AA follow a similar trajectory, underscoring that shared mechanisms drive disease in both murine and human AA. Our study represents a comprehensive, systematic interrogation of lymphocyte heterogeneity in AA and uncovers a novel framework for AA-associated CD8+ T cells with implications for the design of future therapeutics.
Keywords: T cells; alopecia areata; autoimmunity; hair follicle; single-cell RNA sequencing.
Conflict of interest statement
N.A. is a scientific advisor and an equity holder in Shennon Biotechnologies and is a consultant for Janssen, Immunitas, 23 and me, and Cellino Pharmaceuticals. A.M.C. is a consultant/scientific advisor for Almirall S.A., Arcutis Biotherapeutics, Inc., Intrinsic Medicine, Inc., and Pfizer, Inc.; is a shareholder of Aclaris Therapeutics, Inc. and Intrinsic Medicine, Inc.; has received research grant support from Pfizer, Inc.; is a coinventor on several patents filed by Columbia University on the use of JAK inhibitors in treating hair loss disorders, which have been licensed to Aclaris Therapeutics, Inc. and sublicensed from Aclaris to Eli Lilly & Co., resulting in intellectual property payments to the coinventors and to Columbia University; serves on the scientific advisory boards for the Dystrophic EB Research Association of America and the National AA Foundation; and is currently President of the American Hair Research Society.
Figures
References
Publication types
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
