

ResR/McdR-regulated protein translation machinery contributes to drug resilience in Mycobacterium tuberculosis
- PMID: 37433855
- PMCID: PMC10336103
- DOI: 10.1038/s42003-023-05059-8
ResR/McdR-regulated protein translation machinery contributes to drug resilience in Mycobacterium tuberculosis
Abstract
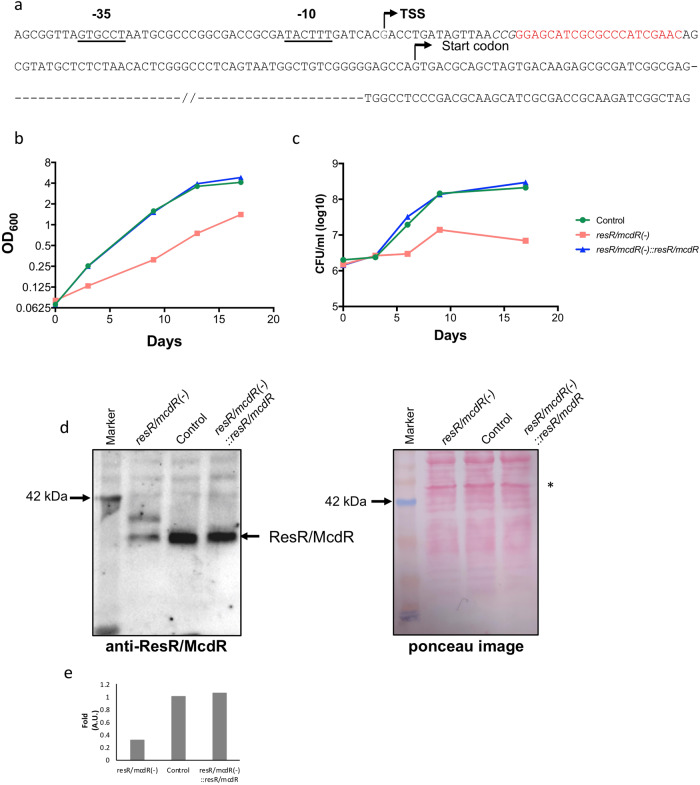
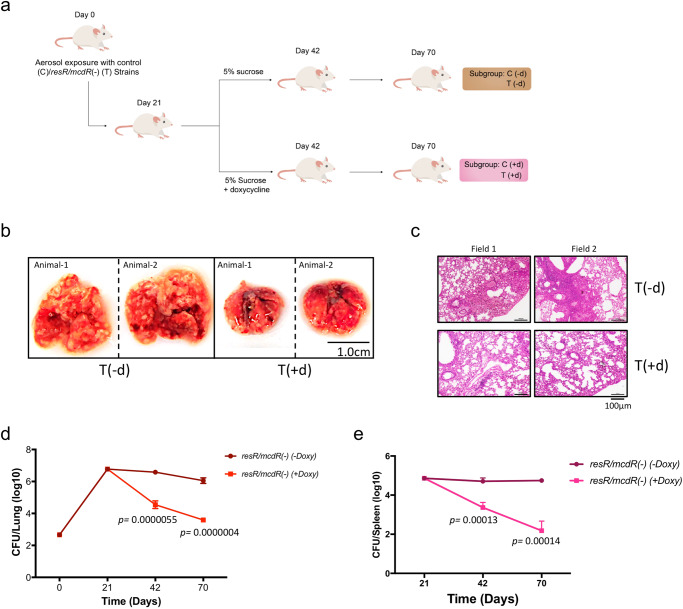
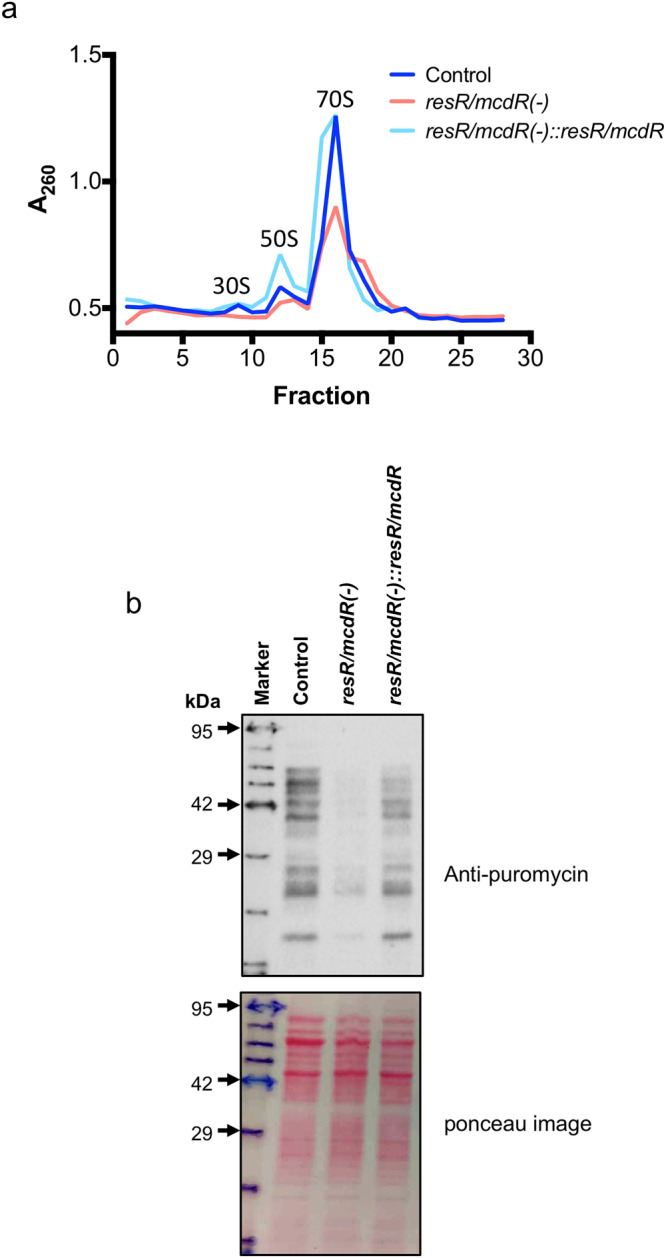
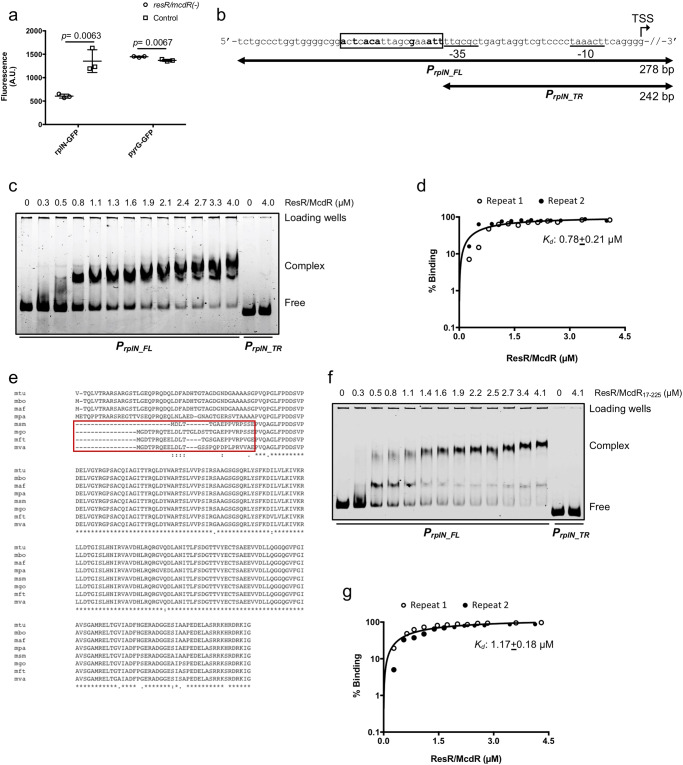
Survival response of the human tuberculosis pathogen, Mycobacterium tuberculosis (Mtb) to a diverse environmental cues is governed through its versatile transcription regulatory mechanisms with the help of a large pool of transcription regulators (TRs). Rv1830 is one such conserved TR, which remains uncharacterized in Mtb. It was named as McdR based on an effect on cell division upon its overexpression in Mycobacterium smegmatis. Recently, it has been implicated in antibiotic resilience in Mtb and reannotated as ResR. While Rv1830 affects cell division by modulating the expression of M. smegmatis whiB2, the underlying cause of its essentiality and regulation of drug resilience in Mtb is yet to be deciphered. Here we show that ResR/McdR, encoded by ERDMAN_2020 in virulent Mtb Erdman, is pivotal for bacterial proliferation and crucial metabolic activities. Importantly, ResR/McdR directly regulates ribosomal gene expression and protein synthesis, requiring distinct disordered N-terminal sequence. Compared to control, bacteria depleted with resR/mcdR exhibit delayed recovery post-antibiotic treatment. A similar effect upon knockdown of rplN operon genes further implicates ResR/McdR-regulated protein translation machinery in attributing drug resilience in Mtb. Overall, findings from this study suggest that chemical inhibitors of ResR/McdR may be proven effective as adjunctive therapy for shortening the duration of TB treatment.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
A Feedback Regulatory Loop Containing McdR and WhiB2 Controls Cell Division and DNA Repair in Mycobacteria.mBio. 2022 Apr 26;13(2):e0334321. doi: 10.1128/mbio.03343-21. Epub 2022 Mar 31. mBio. 2022. PMID: 35357209 Free PMC article.
-
Tuberculosis treatment failure associated with evolution of antibiotic resilience.Science. 2022 Dec 9;378(6624):1111-1118. doi: 10.1126/science.abq2787. Epub 2022 Dec 8. Science. 2022. PMID: 36480634 Free PMC article.
-
Extra-ribosomal functions of Mtb RpsB in imparting stress resilience and drug tolerance to mycobacteria.Biochimie. 2020 Oct;177:87-97. doi: 10.1016/j.biochi.2020.08.007. Epub 2020 Aug 21. Biochimie. 2020. PMID: 32828823
-
[Development of antituberculous drugs: current status and future prospects].Kekkaku. 2006 Dec;81(12):753-74. Kekkaku. 2006. PMID: 17240921 Review. Japanese.
-
Protein synthesis in Mycobacterium tuberculosis as a potential target for therapeutic interventions.Mol Aspects Med. 2021 Oct;81:101002. doi: 10.1016/j.mam.2021.101002. Epub 2021 Jul 31. Mol Aspects Med. 2021. PMID: 34344520 Review.
References
-
- WHO. Global Tuberculosis Report 2021. (2021).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
