

LRP8-mediated selenocysteine uptake is a targetable vulnerability in MYCN-amplified neuroblastoma
- PMID: 37435859
- PMCID: PMC10405063
- DOI: 10.15252/emmm.202318014
LRP8-mediated selenocysteine uptake is a targetable vulnerability in MYCN-amplified neuroblastoma
Abstract
Ferroptosis has emerged as an attractive strategy in cancer therapy. Understanding the operational networks regulating ferroptosis may unravel vulnerabilities that could be harnessed for therapeutic benefit. Using CRISPR-activation screens in ferroptosis hypersensitive cells, we identify the selenoprotein P (SELENOP) receptor, LRP8, as a key determinant protecting MYCN-amplified neuroblastoma cells from ferroptosis. Genetic deletion of LRP8 leads to ferroptosis as a result of an insufficient supply of selenocysteine, which is required for the translation of the antiferroptotic selenoprotein GPX4. This dependency is caused by low expression of alternative selenium uptake pathways such as system Xc- . The identification of LRP8 as a specific vulnerability of MYCN-amplified neuroblastoma cells was confirmed in constitutive and inducible LRP8 knockout orthotopic xenografts. These findings disclose a yet-unaccounted mechanism of selective ferroptosis induction that might be explored as a therapeutic strategy for high-risk neuroblastoma and potentially other MYCN-amplified entities.
Keywords: ferroptosis; neuroblastoma; selenocysteine; selenoprotein; synthetic lethality.
© 2023 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare the following financial interests/personal relationships, which may be considered as potential competing interests: MC is the cofounder of ROSCUE THERAPEUTICS GmbH and author of a patent application related to ferroptosis. GS‐F is coauthor of patent applications related to SLCs, cofounder of Solgate GmbH, and the Academic Project Coordinator of the IMI grants RESOLUTE and Resolution in partnership with Pfizer, Novartis, Bayer, Sanofi, Boehringer‐Ingelheim and Vifor Pharma. The GS‐F laboratory receives funds from Pfizer. All other authors declare no other relevant conflicts of interest.
Figures
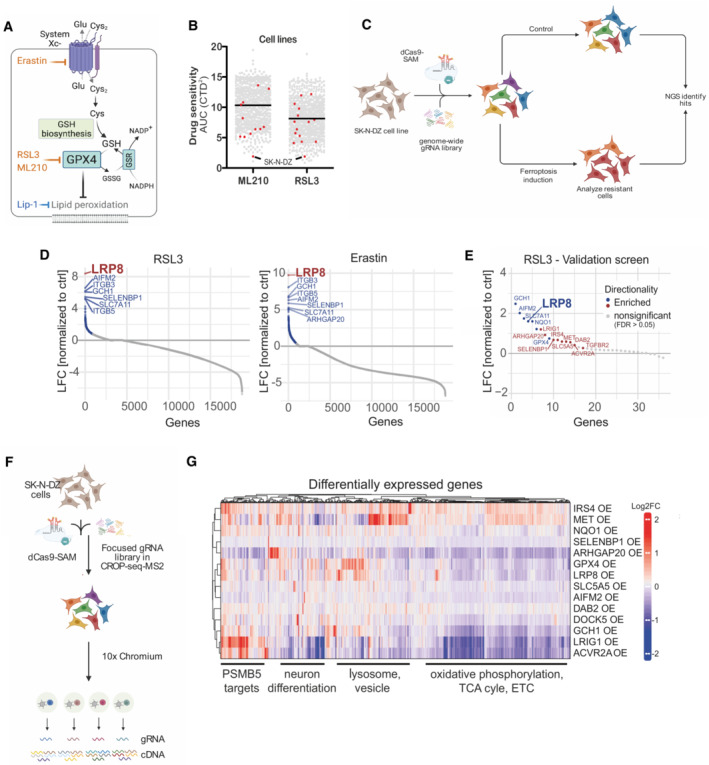
Schematic depiction of ferroptosis regulators and small molecule modulators. Inhibitors are shown in light blue, and inducers in orange.
Analyzis of the depmap portal (
www.depmap.org ) reveals MYCN‐amplified SK‐N‐DZ as hypersensitive cell lines to the GPX4 inhibitors RSL3 and ML210.Strategy of the genome‐wide CRISPR activation (CRISPRa) screen in MYCN‐amplified SK‐N‐DZ cells.
Overexpression phenotypes conferring resistance to 300 nM RSL3 (left) or 1 μM Erastin (right) treatment. Significant hits are marked in blue (FDR ≤ 0.05), while the highest‐scoring hit, LRP8, is highlighted in red.
Overexpression phenotypes conferring resistance to RSL3 (100 nM) induced ferroptosis in the pooled validation CRISPRa screen. Significantly enriched hits (FDR ≤ 0.05) are marked and labeled in blue (known ferroptosis regulators) or red (novel regulators identified in this study). The highest‐scoring hit from the primary screens, LRP8, is highlighted.
Strategy of the single‐cell CRISPRa screen to characterize hits from the ferroptosis‐resistance screen. Guide RNA labels are recovered alongside the whole transcriptome readout for each cell.
Transcriptomic consequences of CRISPRa of 14 scoring hits from the primary and the validation screens. Each row represents one CRISPRa cluster. For each cluster, the top 50 genes with the most significant differential expression (compared to the nontargeting control cluster) were selected and merged to a signature gene list represented by the columns. Columns and rows were hierarchically clustered based on Pearson's correlation.
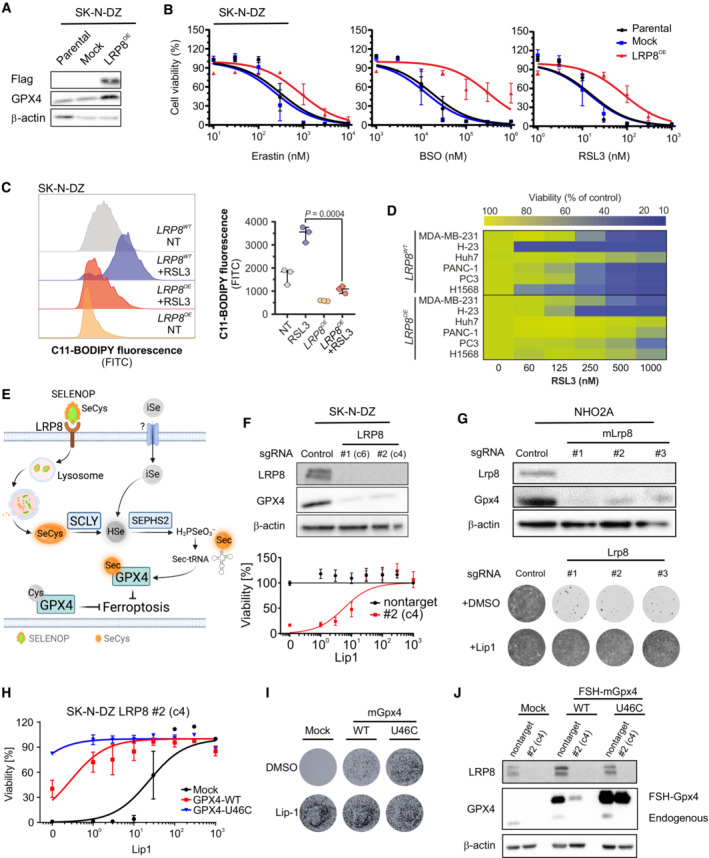
Immunoblot analyzis of FLAG and GPX4 in SK‐N‐DZ cells overexpressing an empty vector or hLRP8‐Flag.
Dose‐dependent toxicity of the ferroptosis inducers Erastin, BSO, and RSL3 in SK‐N‐DZ cell lines stably transduced with a vector expressing hLRP8‐Flag. Data are the mean ± SEM of n = 3 wells of a 96‐well plate from three independent experiments.
Flow cytometry analyzis of BODIPY 581/591 C11 oxidation in SK‐N‐DZ overexpressing hLRP8‐Flag induced by RSL3 treatment (100 nM, 6 h) Data represent the mean ± SEM; n = 3 samples. Statistical analysis was performed using a two‐tailed Student's t‐test.
Heat map depicting the dose‐dependent response of RSL3 in a panel of cell lines overexpressing hLRP8‐Flag.
Schematic representation of selenium uptake mechanisms.
Generation and characterization of LRP8 knockout cell lines using two independent gRNAs. Upper panel, immunoblot analyzis of LRP8 expression in cells transduced with gRNA targeting LRP8. Lower panel, the viability of cell lines, either wild‐type or knockout for LRP8 in the presence of increasing concentrations of Lip‐1. Data are the mean ± SEM of n = 3 wells of a 96‐well plate from two independent experiments.
Recapitulation of LRP8 dependency in the NHO2A cell line derived from a murine model of MYCN amplification. The upper panel depicts the immunoblot of Lrp8 and Gpx4 in cells expressing three independent gRNAs targeting Lrp8. The lower panel shows the clonogenic capacity of Lrp8‐deficient cells and the protective effect of Lip‐1 (500 nM).
Dose‐dependent response of SK‐N‐DZ LRP8 knockout clonal cell line (#2) expressing wild‐type and U46C‐GPX4 in the present of Lip1. Data are the mean ± SEM of n = 3 wells of a 96‐well plate from two independent experiments.
The clonogenic capacity of an SK‐N‐DZ LRP8 knockout clonal cell line (#2) expressing wild‐type and U46C‐GPX4.
Immunoblot analyzis of FLAG and GPX4 in SK‐N‐DZ cells overexpressing flag‐tagged WT or a U46C variant of GPX4 in wild‐type and knockout LRP8 background.
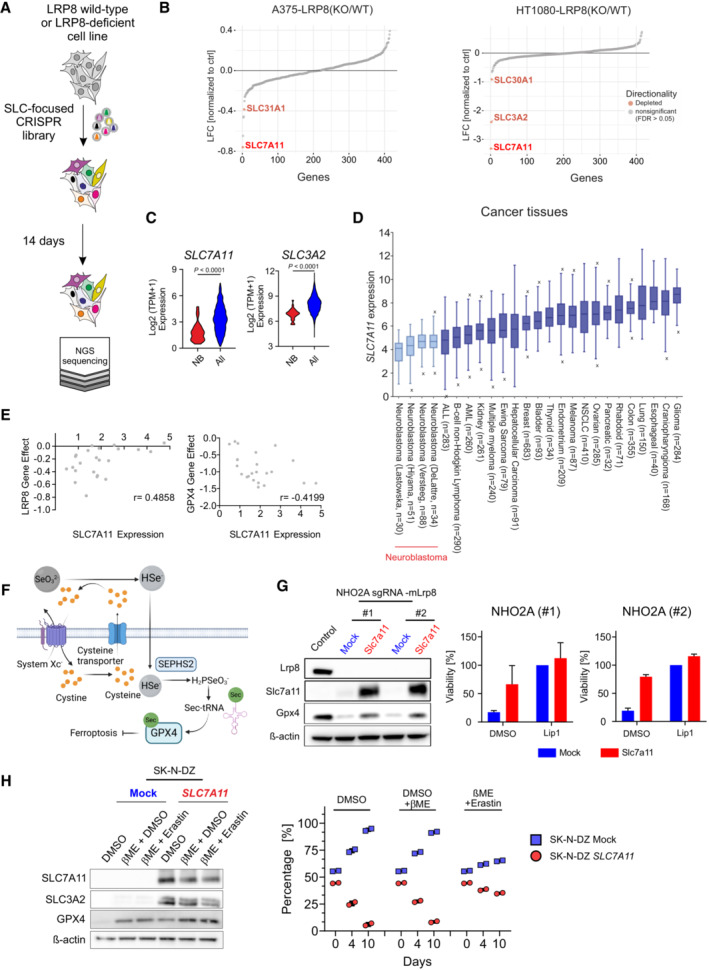
Schematic representation of the SLC‐focused CRISPR knockout screen in the LRP8 wild‐type and LRP8‐deficient cell lines.
Results of a CRISPR deletion screen conducted in H1080 and A375 cell lines displayed a log2 fold change between LRP8‐deficient and wild‐type cells.
Comparison of SLC7A11 expression in a panel of 23 neuroblastoma cell lines against 1,349 non‐neuroblastoma cell lines demonstrating the lineage‐specific lower expression of SLC7A11 and SLC3A2 (
http://www.depmap.org ). Statistical analysis was performed using an Unpaired t‐test. ****P < 0.0001.SLC7A11 expression in cancer tissues of different entities. Data retrieved from the
R2database . The center line in the box plots indicates the median value. Lower and upper hinges represent the 25th and 75th quantiles, the whiskers denote the 1.5× interquartile range, and outliers are marked in the plot.Dot plot depicting the correlation of the dependency of neuroblastoma cell lines on LRP8 and GPX4 (CERES score of −1 means full dependency based on CRISPR–Cas9 knockout screening data) and the expression levels of SLC7A11 in a panel of 27 neuroblastoma cell lines (depmap portal;
https://depmap.org/portal/ ). Cell lines with low expression of SLC7A11 were found to be dependent on LRP8 (Pearson correlation r: 0.4858).Schematic representation of system Xc− of selenium uptake mechanisms.
The immunoblot of Lrp8, Slc7a11 and Gpx4 in NHO2A stably expressing Cas9 and two independent sgRNAs targeting Lrp8 in an SLC7A11 or mock expressing background and viability of the cell lines in the presence of the DMSO, Lip1 (500 nM), or a combination of both. Cell viability assays depicted are mean ± SD of two independently performed experiments (n = 4 wells of a 96‐well plate).
Generation and characterization of cells overexpressing SLC7A11. Immunoblot analysis of SLC7A11 and SLC3A2 from SK‐N‐DZ cells overexpressing SLC7A11 or mock. Cell competition assay of SK‐N‐DZ cell line overexpressing SLC7A11 or mock controls. For the experiment, cells were seeded at a ratio of 50/50 and 50,000 events were measured via flow cytometry at the depicted time points. Rescue experiments were performed in the presence of 50 μM beta‐Mercaptoethanol (beta‐ME) and 2 μM Erastin. Bars display percentage of eGFP‐ (blue) and Scarlet‐cells (red) with the means ± SD of two independent experiments. Statistical analysis was performed using an Unpaired t‐test comparing the abundance of SK‐N‐DZ cells overexpressing SLC7A11 (Red).
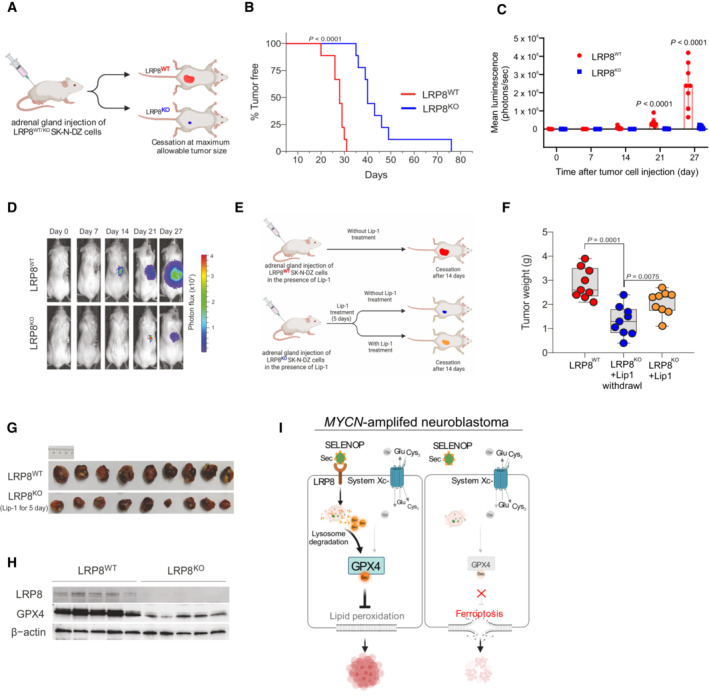
Schematic representation of the orthotopic implantation of control (LRP8WT) or LRP8‐deficient (LRP8KO) SK‐N‐DZ cell lines.
Kaplan–Meier plot displaying tumor‐free survival (TFS) for mice injected orthotopically with LRP8WT (red, n = 9) or LRP8KO (blue, n = 9) SK‐N‐DZ cells. A Gehan–Breslow–Wilcoxon test was conducted for statistical analyzis.
Tumor growth upon orthotopic implantation of LRP8WT (red, n = 9) or LRP8KO (blue, n = 9) of SK‐N‐DZ cell line. Data represent the mean ± SEM; Mann–Whitney test one‐tailed P‐values are indicated.
Representative luminescence images from each group are sown in (C).
Outline of the orthotopic implantation and treatment scheme with Liproxstatin‐1 (Lip‐1) of LRP8WT and LRP8KO SK‐N‐DZ cells.
Tumor weight of orthotopically implanted LRP8WT (red, n = 9) or LRP8KO (blue and yellow, n = 9 each) SK‐N‐DZ cells. All mice were treated with Lip‐1 for 5 days. After this, treatment was ceased for LRP8WT (red) and LRP8KO cohorts (blue) or maintained for an additional 14 days (yellow); see Materials and Methods for details. Tumors were analyzed at the endpoint. The center line in the box plots indicates the median value. Lower and upper hinges represent the 25th and 75th quantiles, the whiskers denote the Max and Min value. Mann–Whitney test one‐tailed P values are indicated.
Representative images of tumors in the control (red) and LRP8KO groups (blue) treated with Lip‐1 for 5 days after implantation.
Immunoblot analyzis of LRP8 and GPX4 levels from orthotopic tumors of LRP8WT or LRP8KO SK‐N‐DZ cells, treated with Lip‐1 for 5 days after implantation, followed by 14 days after randomization.
Schematic representation for the proposed model of LRP8 inhibition essentiality. Comparison of selenium /selenocysteine uptake mechanisms via LRP8 and system Xc− in proliferating MYCN‐amplified cells, depicting the contribution of primarily LRP8/SELENOP supporting selenoprotein translation (left panel). Inhibition of LRP8 in system Xc− low cell selectively triggers ferroptosis in MYCN‐amplified neuroblastoma (right panel). SELENOP, Selenoprotein P; iSe, inorganic selenium; SeCys, Selenocysteine.
References
-
- Brutsch SH, Wang CC, Li L, Stender H, Neziroglu N, Richter C, Kuhn H, Borchert A (2015) Expression of inactive glutathione peroxidase 4 leads to embryonic lethality, and inactivation of the Alox15 gene does not rescue such knock‐in mice. Antioxid Redox Signal 22: 281–293 - PubMed
-
- Burk RF, Hill KE (2015) Regulation of selenium metabolism and transport. Annu Rev Nutr 35: 109–134 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
