

Models of KPTN-related disorder implicate mTOR signalling in cognitive and overgrowth phenotypes
- PMID: 37437211
- PMCID: PMC10629792
- DOI: 10.1093/brain/awad231
Models of KPTN-related disorder implicate mTOR signalling in cognitive and overgrowth phenotypes
Abstract
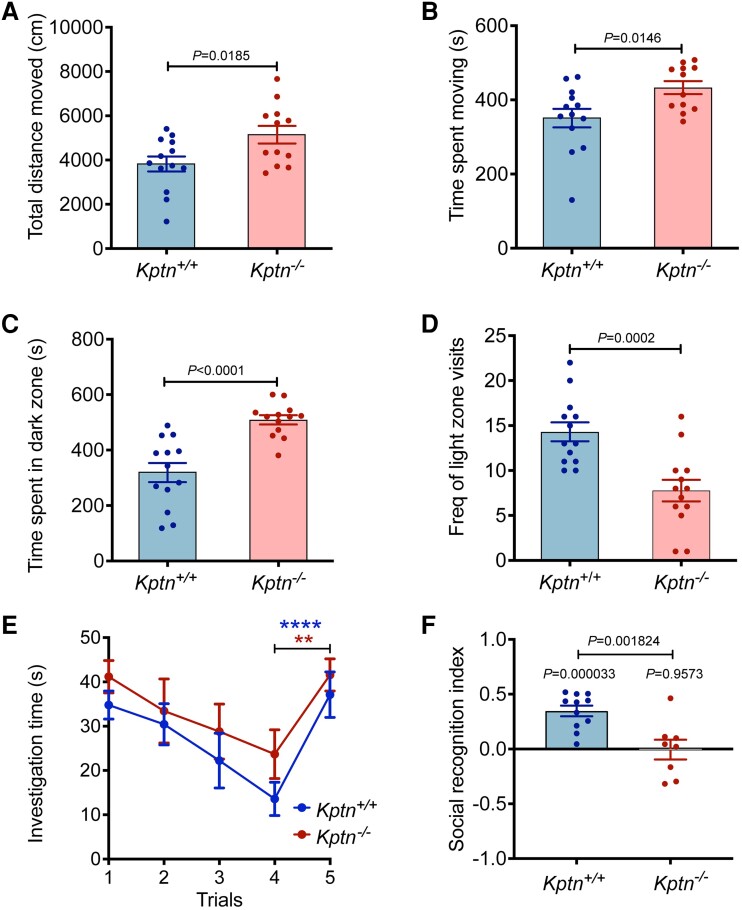
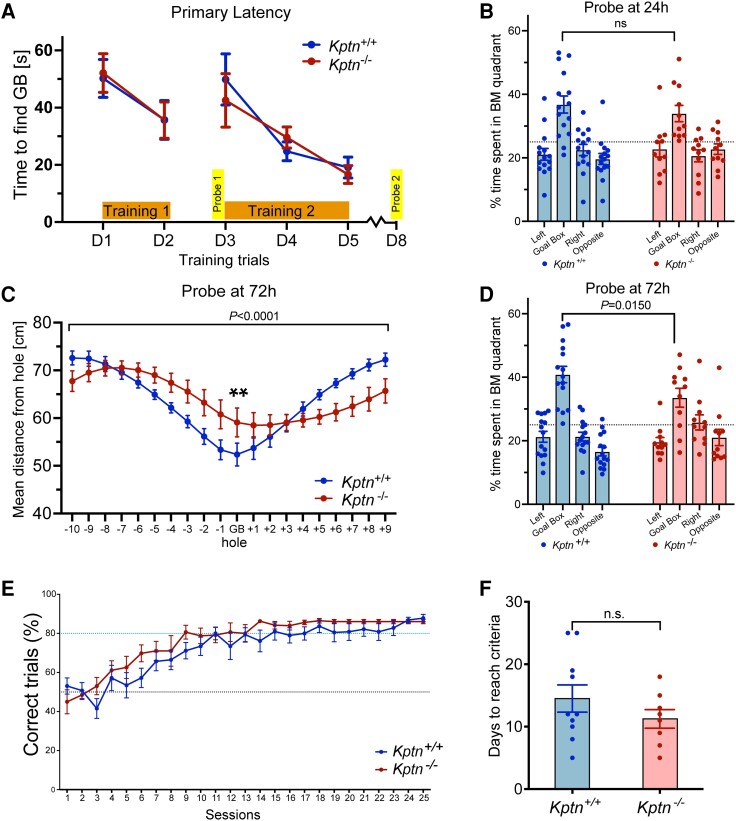
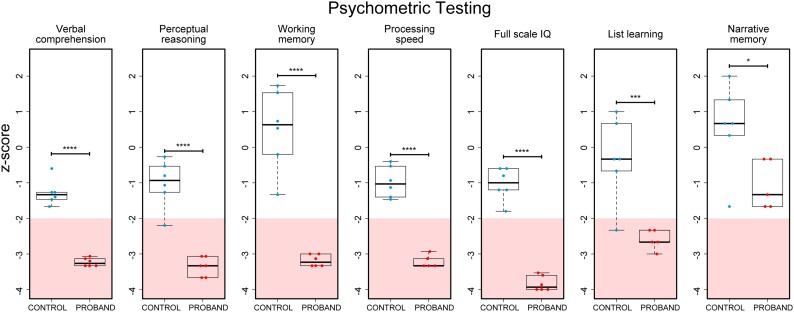
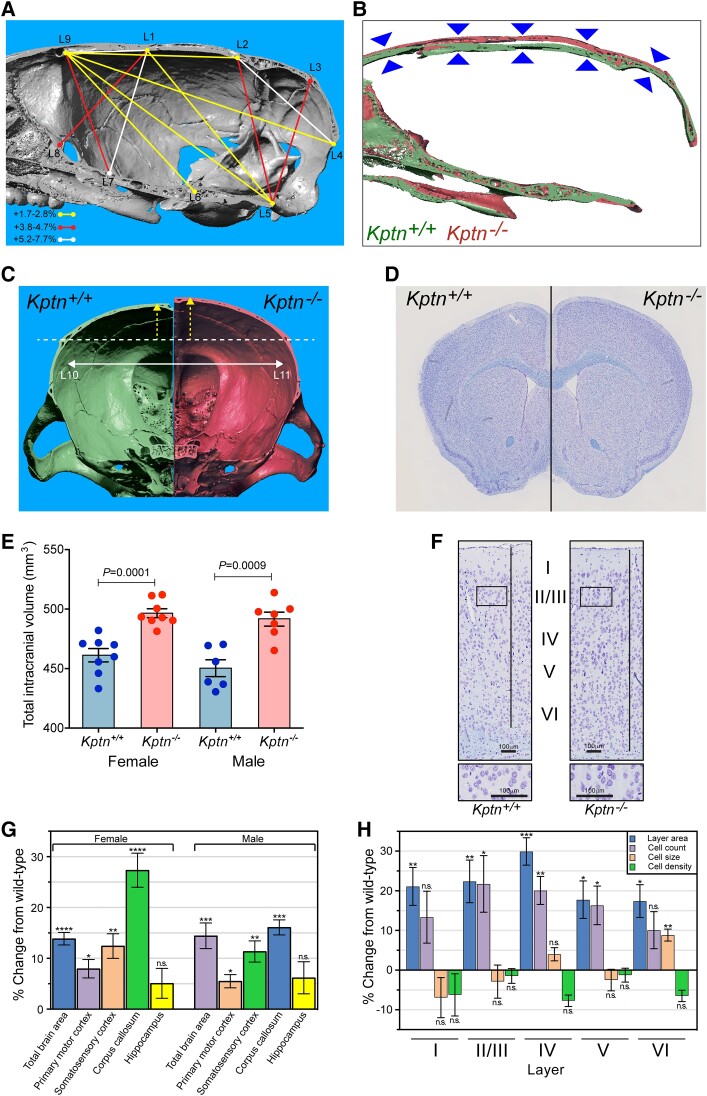
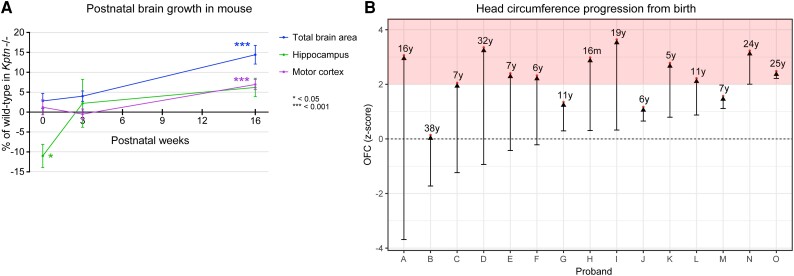
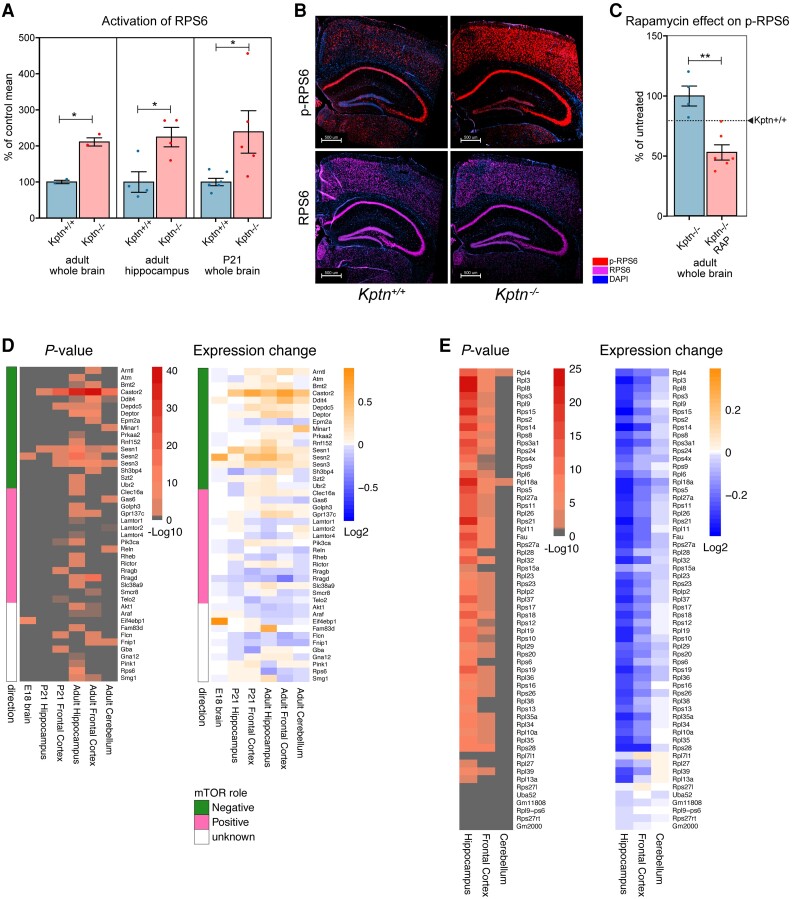
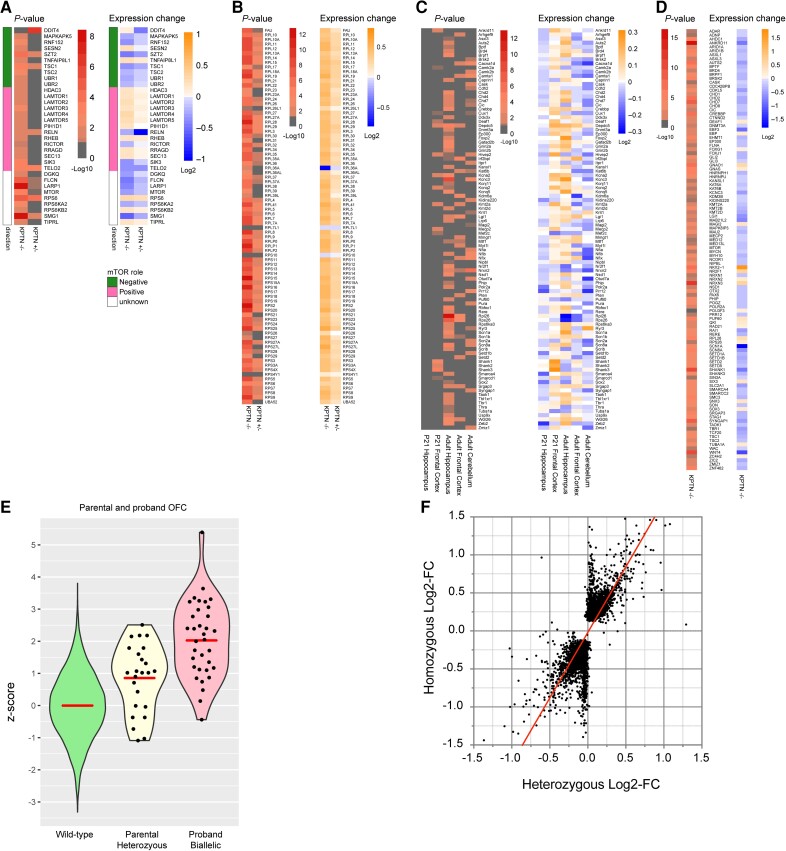
KPTN-related disorder is an autosomal recessive disorder associated with germline variants in KPTN (previously known as kaptin), a component of the mTOR regulatory complex KICSTOR. To gain further insights into the pathogenesis of KPTN-related disorder, we analysed mouse knockout and human stem cell KPTN loss-of-function models. Kptn -/- mice display many of the key KPTN-related disorder phenotypes, including brain overgrowth, behavioural abnormalities, and cognitive deficits. By assessment of affected individuals, we have identified widespread cognitive deficits (n = 6) and postnatal onset of brain overgrowth (n = 19). By analysing head size data from their parents (n = 24), we have identified a previously unrecognized KPTN dosage-sensitivity, resulting in increased head circumference in heterozygous carriers of pathogenic KPTN variants. Molecular and structural analysis of Kptn-/- mice revealed pathological changes, including differences in brain size, shape and cell numbers primarily due to abnormal postnatal brain development. Both the mouse and differentiated induced pluripotent stem cell models of the disorder display transcriptional and biochemical evidence for altered mTOR pathway signalling, supporting the role of KPTN in regulating mTORC1. By treatment in our KPTN mouse model, we found that the increased mTOR signalling downstream of KPTN is rapamycin sensitive, highlighting possible therapeutic avenues with currently available mTOR inhibitors. These findings place KPTN-related disorder in the broader group of mTORC1-related disorders affecting brain structure, cognitive function and network integrity.
Keywords: animal model; iPSC; mTOR; macrocephaly; neurodevelopmental disorders; recessive.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Guarantors of Brain.
Conflict of interest statement
M.E.H. is a co-founder, shareholder, non-executive director of Congenica, and an advisor to Astra-Zeneca.
Figures
Comment in
-
Growth signalling pathways in brain size determination and neurodevelopmental disorders.Brain. 2023 Nov 2;146(11):4399-4400. doi: 10.1093/brain/awad365. Brain. 2023. PMID: 37934918 No abstract available.
References
-
- Azevedo FAC, Carvalho LRB, Grinberg LT, et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J Comp Neurol. 2009;513:532–541. - PubMed
-
- Zoghbi HY. Postnatal neurodevelopmental disorders: Meeting at the synapse? Science. 2003;302:826–830. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
