

External globus pallidus input to the dorsal striatum regulates habitual seeking behavior in male mice
- PMID: 37438336
- PMCID: PMC10338526
- DOI: 10.1038/s41467-023-39545-8
External globus pallidus input to the dorsal striatum regulates habitual seeking behavior in male mice
Abstract
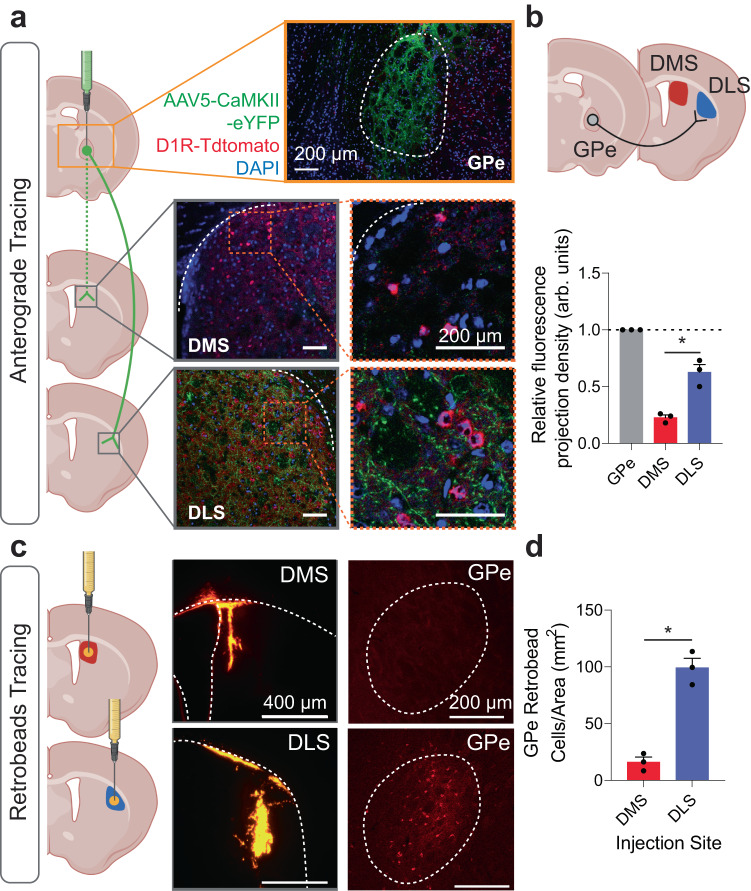
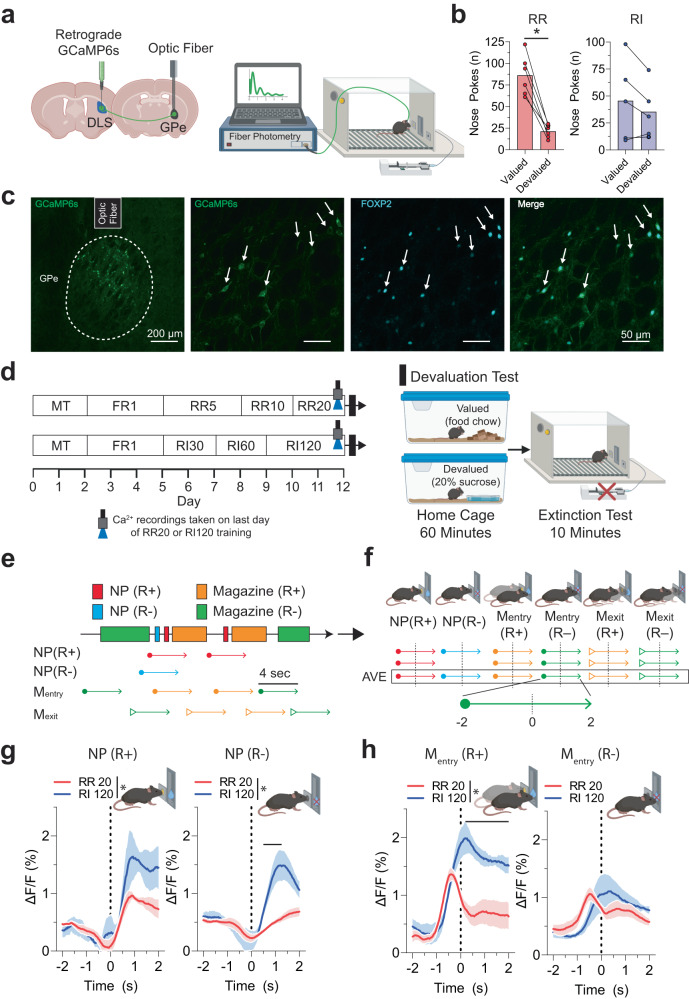
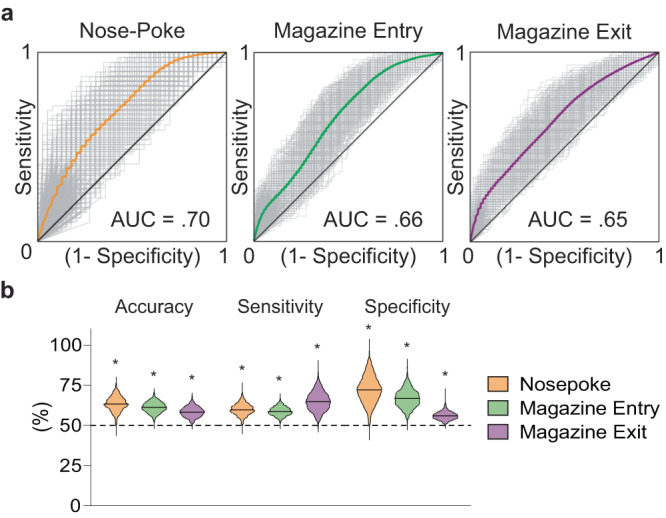
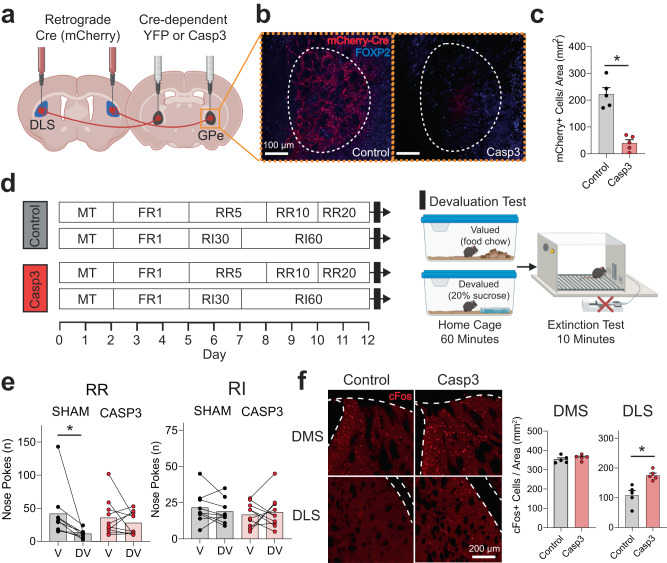
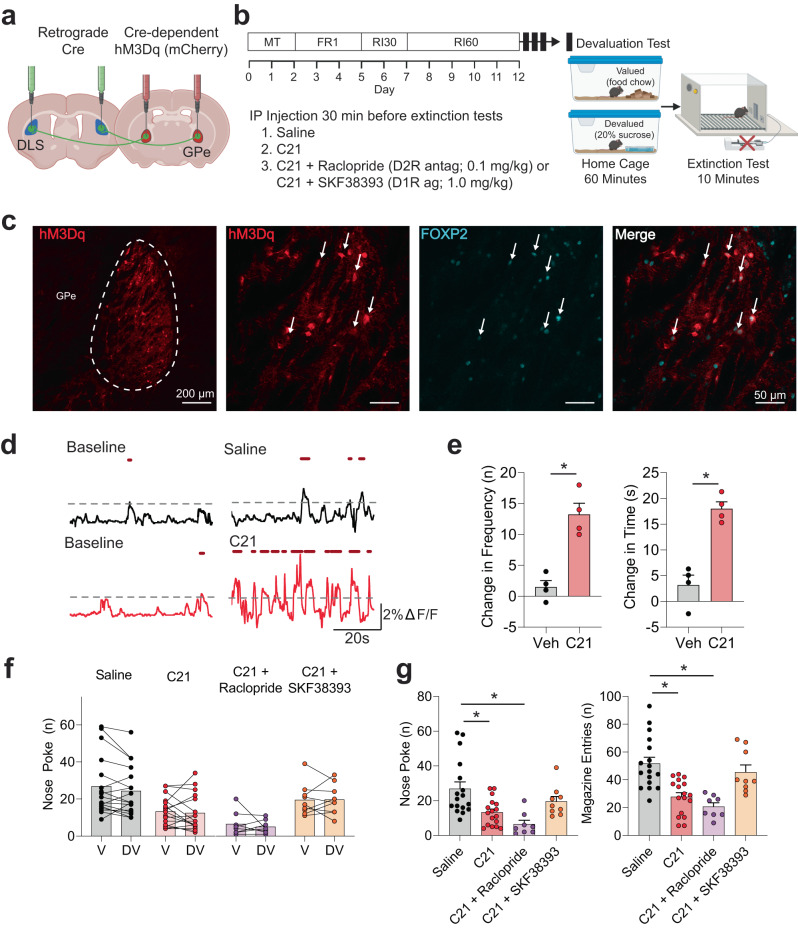
The external globus pallidus (GPe) coordinates action-selection through GABAergic projections throughout the basal ganglia. GPe arkypallidal (arky) neurons project exclusively to the dorsal striatum, which regulates goal-directed and habitual seeking. However, the role of GPe arky neurons in reward-seeking remains unknown. Here, we identified that a majority of arky neurons target the dorsolateral striatum (DLS). Using fiber photometry, we found that arky activities were higher during random interval (RI; habit) compared to random ratio (RR; goal) operant conditioning. Support vector machine analysis demonstrated that arky neuron activities have sufficient information to distinguish between RR and RI behavior. Genetic ablation of this arkyGPe→DLS circuit facilitated a shift from goal-directed to habitual behavior. Conversely, chemogenetic activation globally reduced seeking behaviors, which was blocked by systemic D1R agonism. Our findings reveal a role of this arkyGPe→DLS circuit in constraining habitual seeking in male mice, which is relevant to addictive behaviors and other compulsive disorders.
© 2023. The Author(s).
Conflict of interest statement
D.S.C. is a scientific advisory board member to Peptron Inc. Peptron had no role in the preparation, review, or approval of the manuscript; nor the decision to submit the manuscript for publication. The remaining authors declare no competing interests.
Figures
Comment in
-
Arky neurons and reward seeking.Nat Neurosci. 2023 Aug;26(8):1319. doi: 10.1038/s41593-023-01412-6. Nat Neurosci. 2023. PMID: 37537348 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
