

CRAT links cholesterol metabolism to innate immune responses in the heart
- PMID: 37443356
- PMCID: PMC10685850
- DOI: 10.1038/s42255-023-00844-5
CRAT links cholesterol metabolism to innate immune responses in the heart
Abstract
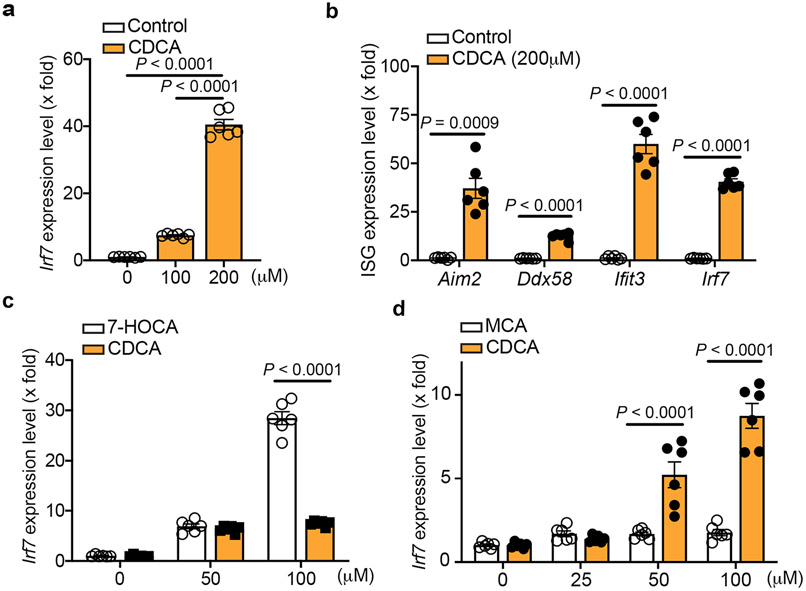
Chronic inflammation is associated with increased risk and poor prognosis of heart failure; however, the precise mechanism that provokes sustained inflammation in the failing heart remains elusive. Here we report that depletion of carnitine acetyltransferase (CRAT) promotes cholesterol catabolism through bile acid synthesis pathway in cardiomyocytes. Intracellular accumulation of bile acid or intermediate, 7α-hydroxyl-3-oxo-4-cholestenoic acid, induces mitochondrial DNA stress and triggers cGAS-STING-dependent type I interferon responses. Furthermore, type I interferon responses elicited by CRAT deficiency substantially increase AIM2 expression and AIM2-dependent inflammasome activation. Genetic deletion of cardiomyocyte CRAT in mice of both sexes results in myocardial inflammation and dilated cardiomyopathy, which can be reversed by combined depletion of caspase-1, cGAS or AIM2. Collectively, we identify a mechanism by which cardiac energy metabolism, cholesterol homeostasis and cardiomyocyte-intrinsic innate immune responses are interconnected via a CRAT-mediated bile acid synthesis pathway, which contributes to chronic myocardial inflammation and heart failure progression.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
















Comment in
-
Cholesterol catabolism and bile acid synthesis in cardiomyocytes promote inflammation and heart failure.Nat Rev Cardiol. 2023 Oct;20(10):647. doi: 10.1038/s41569-023-00917-8. Nat Rev Cardiol. 2023. PMID: 37524809 No abstract available.
References
-
- Hunt SA et al. ACC/AHA guidelines for the evaluation and management of chronic heart failure in the adult: executive summary a report of the American College of Cardiology/American Heart Association Task Force on practice guidelines (committee to revise the 1995 guidelines for the evaluation and management of heart failure): developed in collaboration with the International Society for Heart and Lung Transplantation; endorsed by the Heart Failure Society of America. Circulation 104, 2996–3007 (2001). - PubMed
-
- McDonagh TA et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J 42, 3599–3726 (2021). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
