

Lrp1 is essential for lethal Rift Valley fever hepatic disease in mice
- PMID: 37450601
- PMCID: PMC10348670
- DOI: 10.1126/sciadv.adh2264
Lrp1 is essential for lethal Rift Valley fever hepatic disease in mice
Abstract
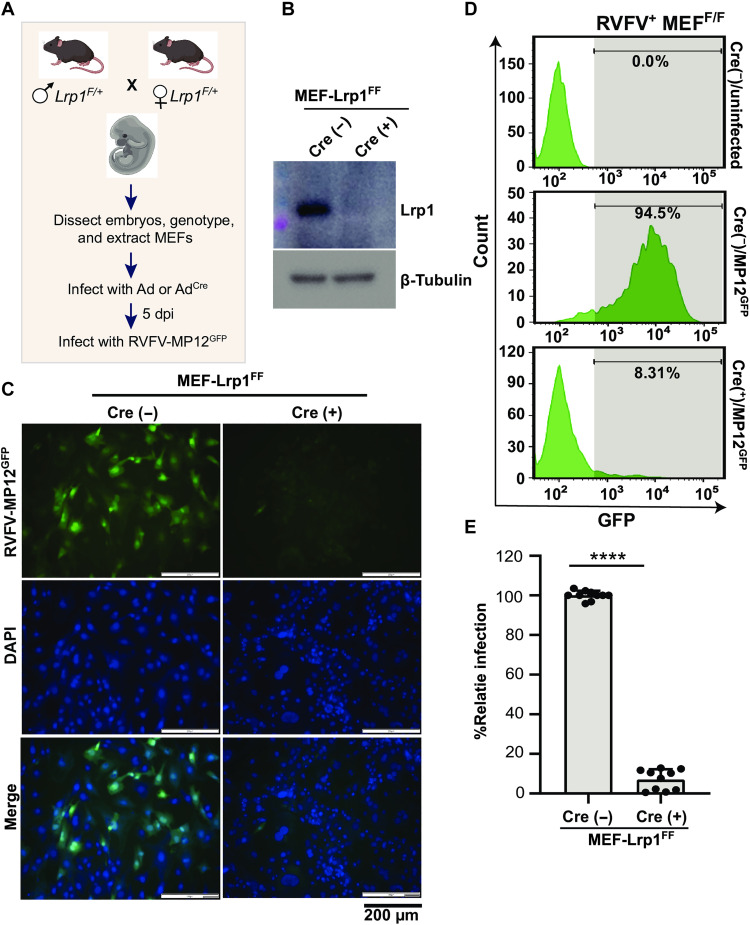
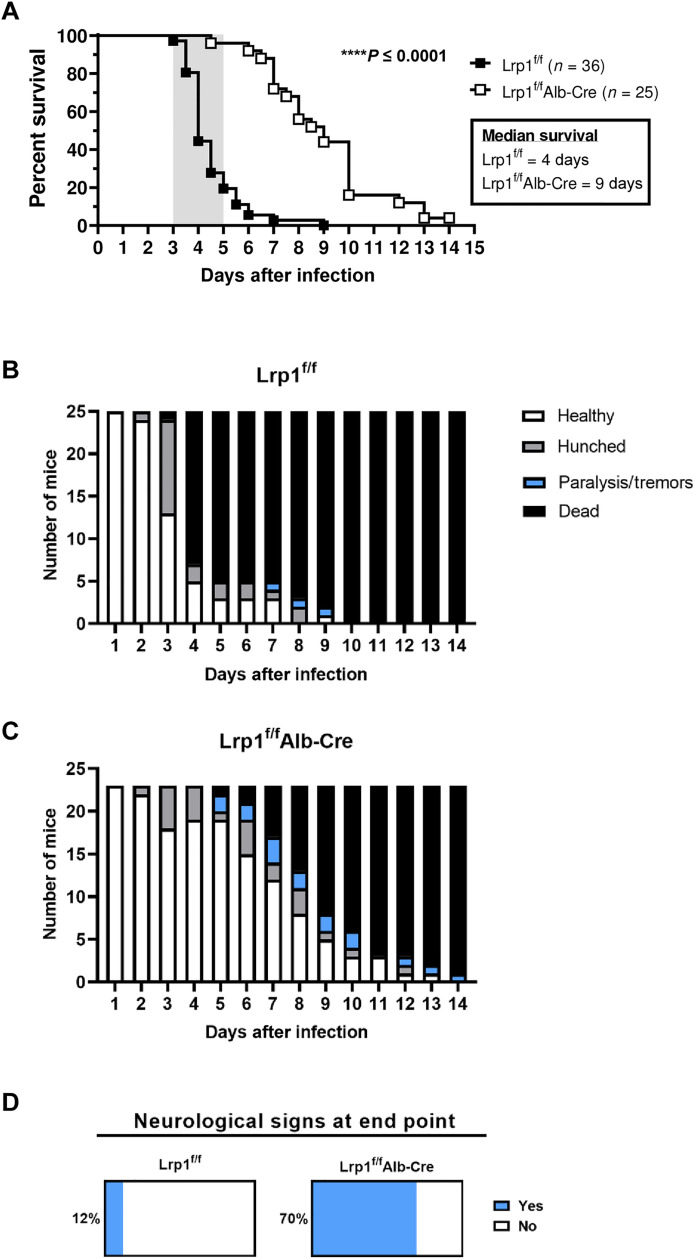
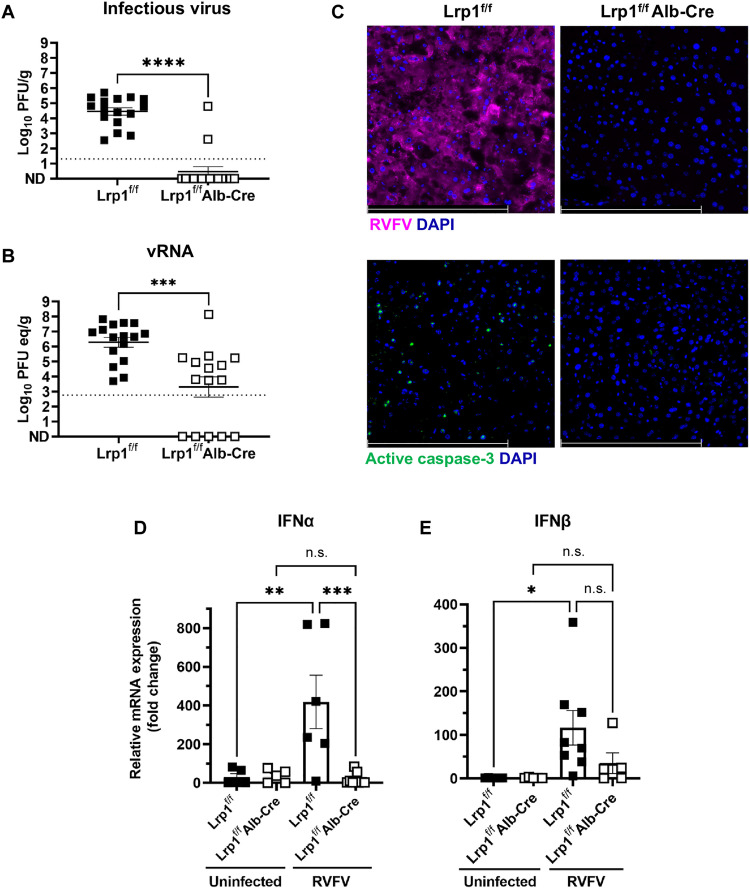
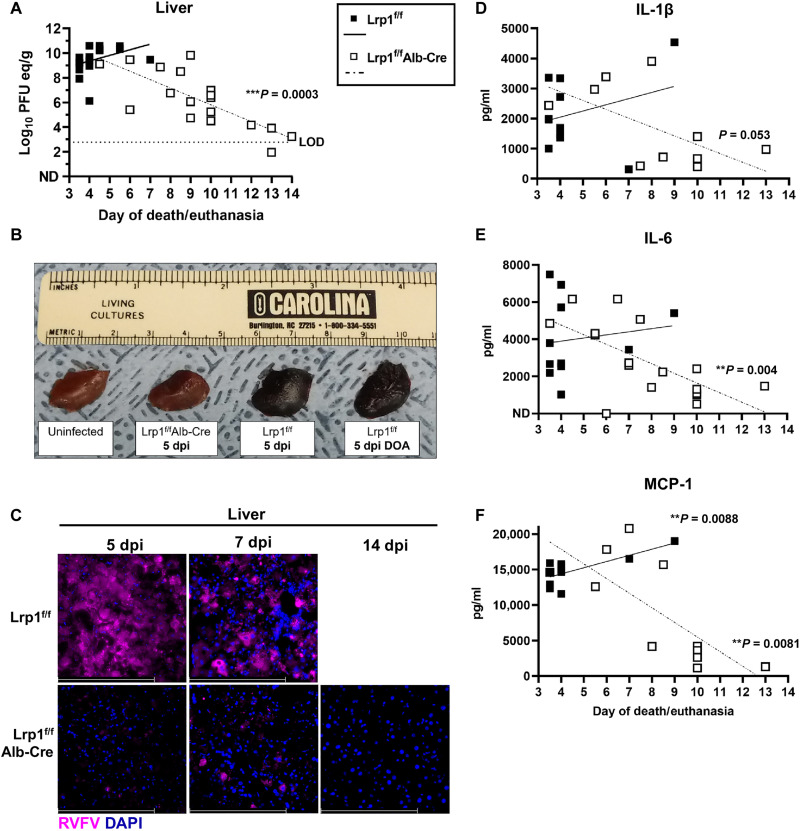
Rift Valley fever virus (RVFV) is an emerging arbovirus found in Africa. While RVFV is pantropic and infects many cells and tissues, viral replication and necrosis within the liver play a critical role in mediating severe disease. The low-density lipoprotein receptor-related protein 1 (Lrp1) is a recently identified host factor for cellular entry and infection by RVFV. The biological significance of Lrp1, including its role in hepatic disease in vivo, however, remains to be determined. Because Lrp1 has a high expression level in hepatocytes, we developed a mouse model in which Lrp1 is specifically deleted in hepatocytes to test how the absence of liver Lrp1 expression affects RVF pathogenesis. Mice lacking Lrp1 expression in hepatocytes showed minimal RVFV replication in the liver, longer time to death, and altered clinical signs toward neurological disease. In contrast, RVFV infection levels in other tissues showed no difference between the two genotypes. Therefore, Lrp1 is essential for RVF hepatic disease in mice.
Figures
References
-
- R. Métras, W. J. Edmunds, C. Youssouffi, L. Dommergues, G. Fournié, A. Camacho, S. Funk, E. Cardinale, G. le Godais, S. Combo, L. Filleul, H. Youssouf, M. Subiros, Estimation of rift valley fever virus spillover to humans during the Mayotte 2018-2019 epidemic. Proc. Natl. Acad. Sci. U.S.A. 117, 24567–24574 (2020). - PMC - PubMed
-
- L. W. Laughlin, J. M. Meegan, L. J. Strausbaugh, D. M. Morens, R. H. Watten, Epidemic rift valley fever in Egypt: Observations of the spectrum of human illness. Trans. R. Soc. Trop. Med. Hyg. 73, 630–633 (1979). - PubMed
-
- M. Al-Hazmi, E. A. Ayoola, M. Abdurahman, S. Banzal, J. Ashraf, A. El-Bushra, A. Hazmi, M. Abdullah, H. Abbo, A. Elamin, E.-T. Al-Sammani, M. Gadour, C. Menon, M. Hamza, I. Rahim, M. Hafez, M. Jambavalikar, H. Arishi, A. Aqeel, Epidemic rift valley fever in Saudi Arabia: A clinical study of severe illness in humans. Clin. Infect. Dis. 36, 245–252 (2003). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
