

pH-dependence of the Plasmodium falciparum chloroquine resistance transporter is linked to the transport cycle
- PMID: 37454114
- PMCID: PMC10349806
- DOI: 10.1038/s41467-023-39969-2
pH-dependence of the Plasmodium falciparum chloroquine resistance transporter is linked to the transport cycle
Abstract
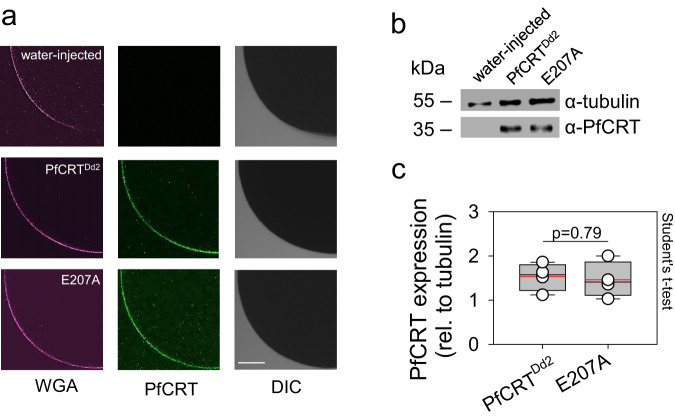
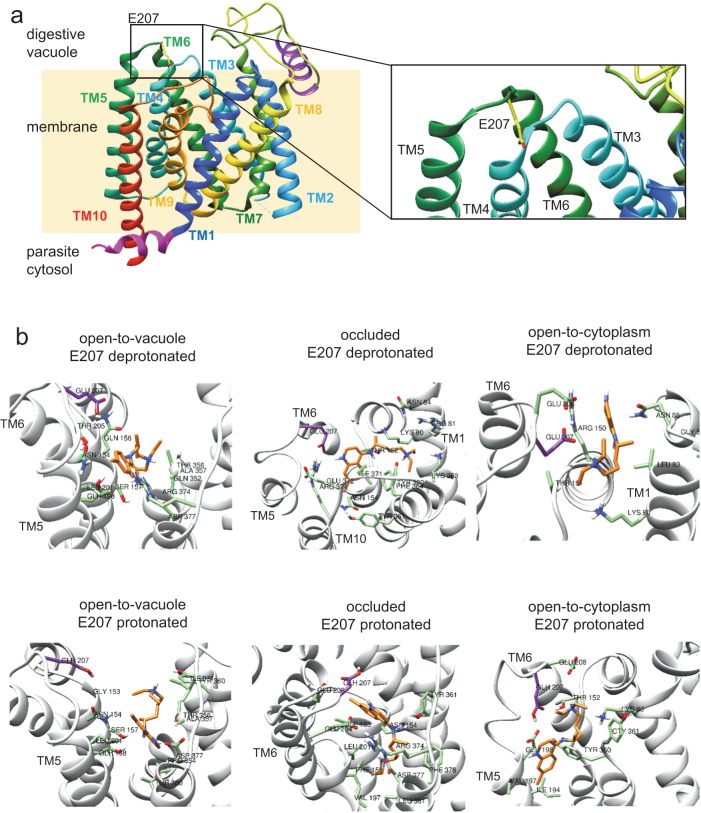
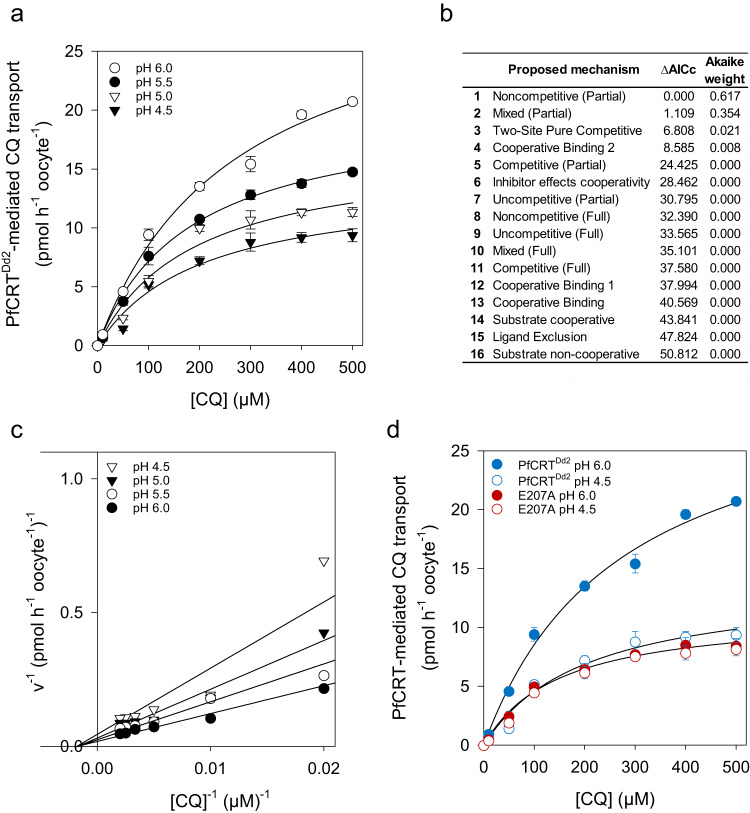
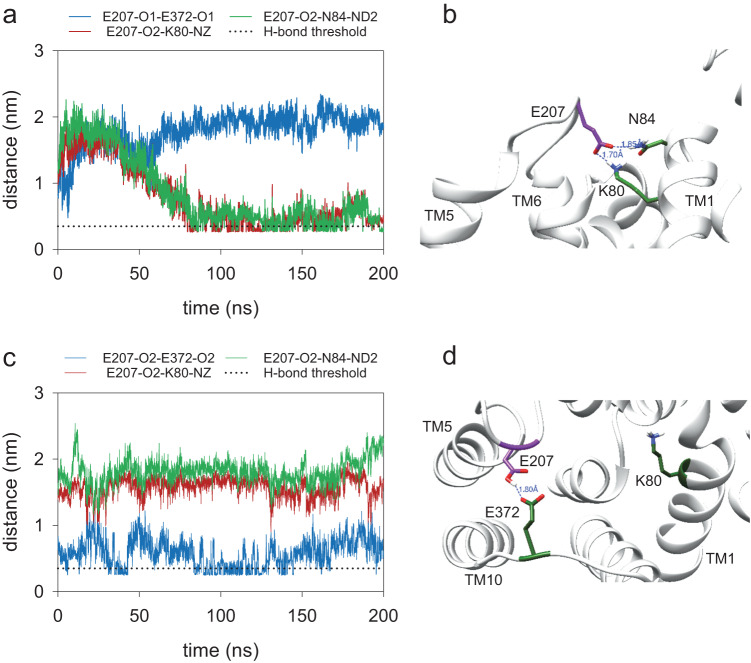
The chloroquine resistance transporter, PfCRT, of the human malaria parasite Plasmodium falciparum is sensitive to acidic pH. Consequently, PfCRT operates at 60% of its maximal drug transport activity at the pH of 5.2 of the digestive vacuole, a proteolytic organelle from which PfCRT expels drugs interfering with heme detoxification. Here we show by alanine-scanning mutagenesis that E207 is critical for pH sensing. The E207A mutation abrogates pH-sensitivity, while preserving drug substrate specificity. Substituting E207 with Asp or His, but not other amino acids, restores pH-sensitivity. Molecular dynamics simulations and kinetics analyses suggest an allosteric binding model in which PfCRT can accept both protons and chloroquine in a partial noncompetitive manner, with increased proton concentrations decreasing drug transport. Further simulations reveal that E207 relocates from a peripheral to an engaged location during the transport cycle, forming a salt bridge with residue K80. We propose that the ionized carboxyl group of E207 acts as a hydrogen acceptor, facilitating transport cycle progression, with pH sensing as a by-product.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Is PfCRT a channel or a carrier? Two competing models explaining chloroquine resistance in Plasmodium falciparum.Trends Parasitol. 2007 Jul;23(7):332-9. doi: 10.1016/j.pt.2007.04.013. Epub 2007 May 10. Trends Parasitol. 2007. PMID: 17493873 Review.
-
A Variant PfCRT Isoform Can Contribute to Plasmodium falciparum Resistance to the First-Line Partner Drug Piperaquine.mBio. 2017 May 9;8(3):e00303-17. doi: 10.1128/mBio.00303-17. mBio. 2017. PMID: 28487425 Free PMC article.
-
A 2-amino quinoline, 5-(3-(2-(7-chloroquinolin-2-yl)ethenyl)phenyl)-8-dimethylcarbamyl-4,6-dithiaoctanoic acid, interacts with PfMDR1 and inhibits its drug transport in Plasmodium falciparum.Mol Biochem Parasitol. 2014 Jun;195(1):34-42. doi: 10.1016/j.molbiopara.2014.05.006. Epub 2014 Jun 8. Mol Biochem Parasitol. 2014. PMID: 24914817
-
A Redox-Active Fluorescent pH Indicator for Detecting Plasmodium falciparum Strains with Reduced Responsiveness to Quinoline Antimalarial Drugs.ACS Infect Dis. 2017 Feb 10;3(2):119-131. doi: 10.1021/acsinfecdis.5b00141. Epub 2016 Dec 7. ACS Infect Dis. 2017. PMID: 28183182
-
PfCRT and its role in antimalarial drug resistance.Trends Parasitol. 2012 Nov;28(11):504-14. doi: 10.1016/j.pt.2012.08.002. Epub 2012 Sep 25. Trends Parasitol. 2012. PMID: 23020971 Free PMC article. Review.
Cited by
-
Combating Malaria: Targeting the Ubiquitin-Proteasome System to Conquer Drug Resistance.Trop Med Infect Dis. 2025 Apr 3;10(4):94. doi: 10.3390/tropicalmed10040094. Trop Med Infect Dis. 2025. PMID: 40278767 Free PMC article. Review.
-
VPS28 regulates triglyceride synthesis via ubiquitination in bovine mammary epithelial cells.Sci Rep. 2024 Dec 28;14(1):31310. doi: 10.1038/s41598-024-82774-0. Sci Rep. 2024. PMID: 39732879 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
