

Variants in ACTC1 underlie distal arthrogryposis accompanied by congenital heart defects
- PMID: 37457373
- PMCID: PMC10345160
- DOI: 10.1016/j.xhgg.2023.100213
Variants in ACTC1 underlie distal arthrogryposis accompanied by congenital heart defects
Abstract
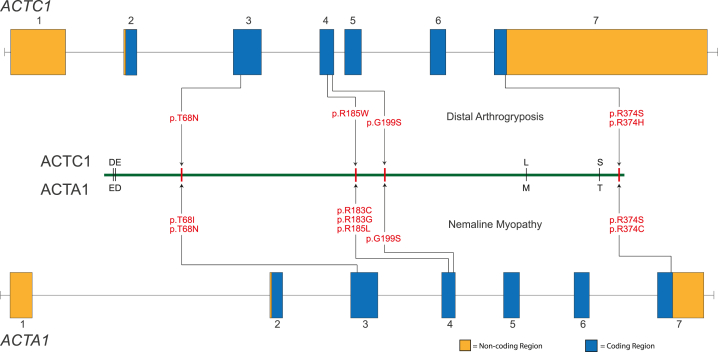
Contraction of the human sarcomere is the result of interactions between myosin cross-bridges and actin filaments. Pathogenic variants in genes such as MYH7, TPM1, and TNNI3 that encode parts of the cardiac sarcomere cause muscle diseases that affect the heart, such as dilated cardiomyopathy and hypertrophic cardiomyopathy. In contrast, pathogenic variants in homologous genes such as MYH2, TPM2, and TNNI2 that encode parts of the skeletal muscle sarcomere cause muscle diseases affecting skeletal muscle, such as distal arthrogryposis (DA) syndromes and skeletal myopathies. To date, there have been few reports of genes (e.g., MYH7) encoding sarcomeric proteins in which the same pathogenic variant affects skeletal and cardiac muscle. Moreover, none of the known genes underlying DA have been found to contain pathogenic variants that also cause cardiac abnormalities. We report five families with DA because of heterozygous missense variants in the gene actin, alpha, cardiac muscle 1 (ACTC1). ACTC1 encodes a highly conserved actin that binds to myosin in cardiac and skeletal muscle. Pathogenic variants in ACTC1 have been found previously to underlie atrial septal defect, dilated cardiomyopathy, hypertrophic cardiomyopathy, and left ventricular noncompaction. Our discovery delineates a new DA condition because of variants in ACTC1 and suggests that some functions of ACTC1 are shared in cardiac and skeletal muscle.
Keywords: Mendelian disease; Mendelian disorder; cardiac defect; cardiomyopathy; congenital contractures; distal arthrogryposis; exome sequencing; gene discovery; molecular dynamics simulations.
© 2023 The Author(s).
Conflict of interest statement
M.J.B. and J.X.C. are the Editor-in-Chief and Deputy Editor of HGG Advances, respectively, and were recused from the editorial handling of this manuscript.
Figures





Update of
-
Variants in ACTC1 underlie distal arthrogryposis accompanied by congenital heart defects.medRxiv [Preprint]. 2023 Mar 9:2023.03.07.23286862. doi: 10.1101/2023.03.07.23286862. medRxiv. 2023. Update in: HGG Adv. 2023 Jun 15;4(3):100213. doi: 10.1016/j.xhgg.2023.100213. PMID: 36945405 Free PMC article. Updated. Preprint.
References
-
- Lindskog C., Linné J., Fagerberg L., Hallström B.M., Sundberg C.J., Lindholm M., Huss M., Kampf C., Choi H., Liem D.A., et al. The human cardiac and skeletal muscle proteomes defined by transcriptomics and antibody-based profiling. BMC Genom. 2015;16:475. doi: 10.1186/s12864-015-1686-y. - DOI - PMC - PubMed
-
- Sung S.S., Brassington A.-M.E., Grannatt K., Rutherford A., Whitby F.G., Krakowiak P.A., Jorde L.B., Carey J.C., Bamshad M. Mutations in Genes Encoding Fast-Twitch Contractile Proteins Cause Distal Arthrogryposis Syndromes. Am. J. Hum. Genet. 2003;72:681–690. doi: 10.1086/368294. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
