

Lysine butyrylation of HSP90 regulated by KAT8 and HDAC11 confers chemoresistance
- PMID: 37460462
- PMCID: PMC10352258
- DOI: 10.1038/s41421-023-00570-y
Lysine butyrylation of HSP90 regulated by KAT8 and HDAC11 confers chemoresistance
Abstract
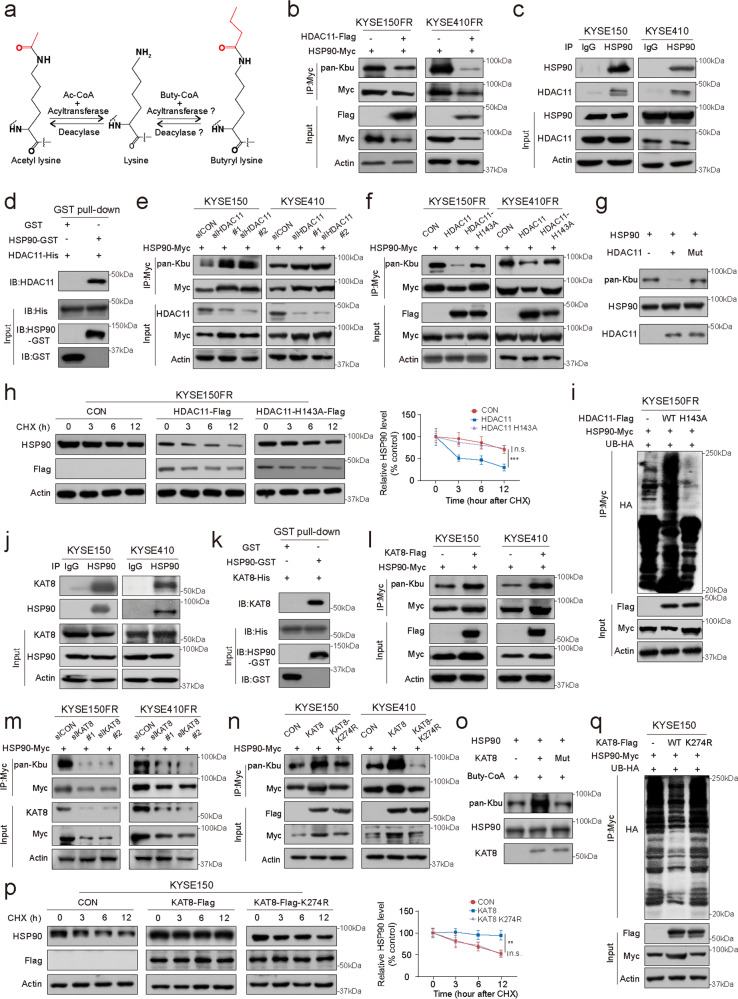
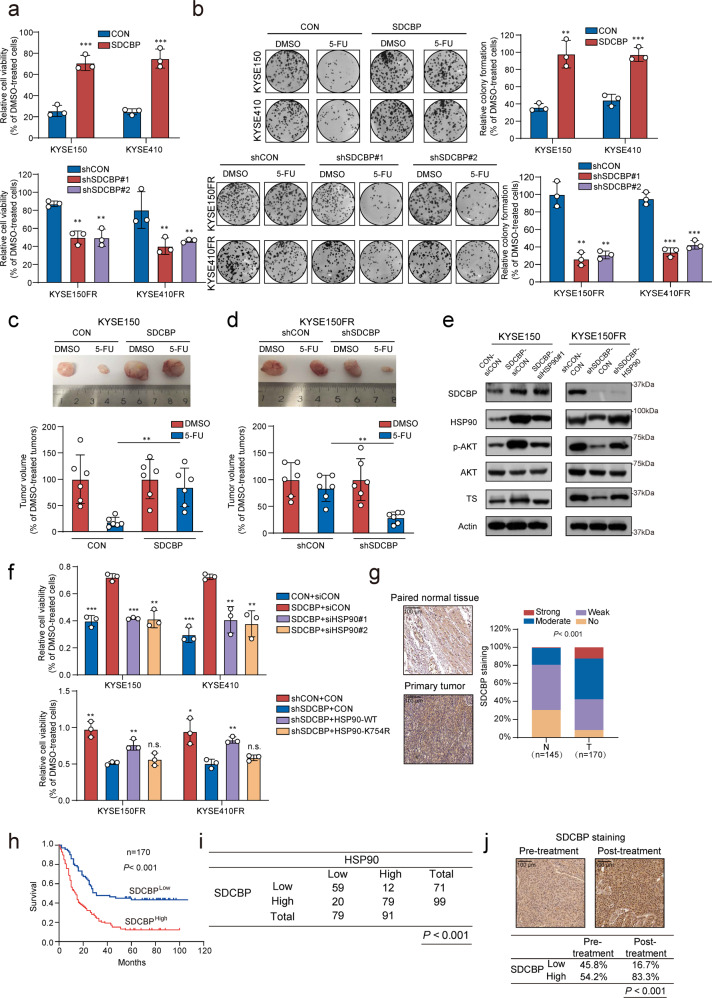
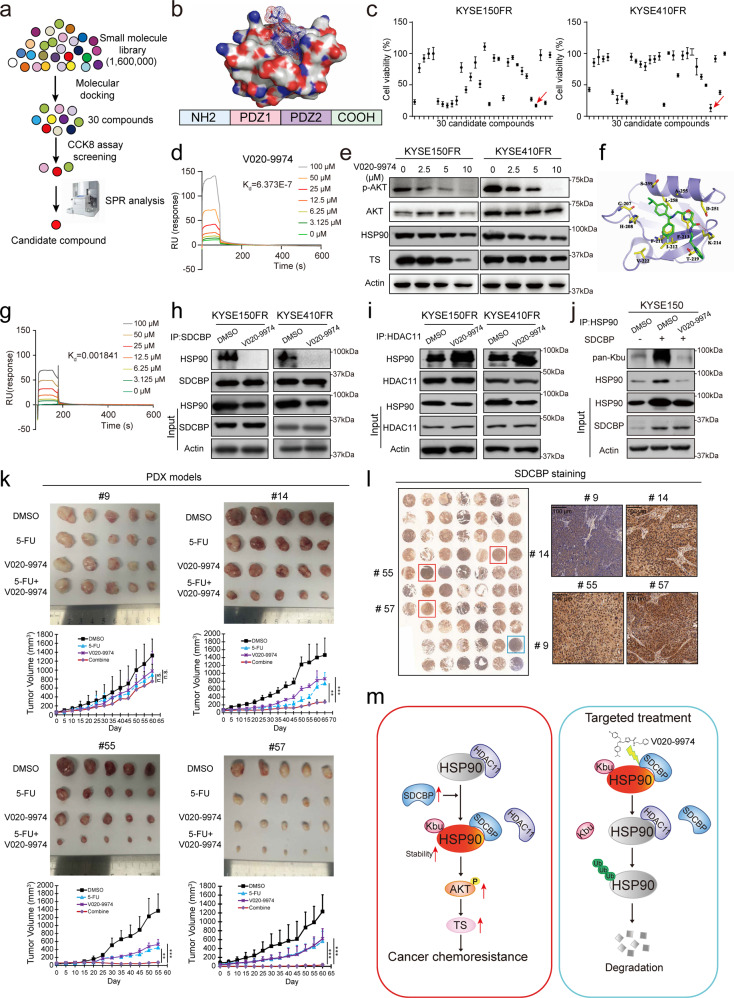
Posttranslational modification dramatically enhances protein complexity, but the function and precise mechanism of novel lysine acylation modifications remain unknown. Chemoresistance remains a daunting challenge to successful treatment. We found that lysine butyrylation (Kbu) is specifically upregulated in chemoresistant tumor cells and tissues. By integrating butyrylome profiling and gain/loss-of-function experiments, lysine 754 in HSP90 (HSP90 K754) was identified as a substrate for Kbu. Kbu modification leads to overexpression of HSP90 in esophageal squamous cell carcinoma (ESCC) and its further increase in relapse samples. Upregulation of HSP90 contributes to 5-FU resistance and can predict poor prognosis in cancer patients. Mechanistically, HSP90 K754 is regulated by the cooperation of KAT8 and HDAC11 as the writer and eraser, respectively; SDCBP increases the Kbu level and stability of HSP90 by binding competitively to HDAC11. Furthermore, SDCBP blockade with the lead compound V020-9974 can target HSP90 K754 to overcome 5-FU resistance, constituting a potential therapeutic strategy.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
