

Fatty acid oxidation organizes mitochondrial supercomplexes to sustain astrocytic ROS and cognition
- PMID: 37460843
- PMCID: PMC10447235
- DOI: 10.1038/s42255-023-00835-6
Fatty acid oxidation organizes mitochondrial supercomplexes to sustain astrocytic ROS and cognition
Abstract
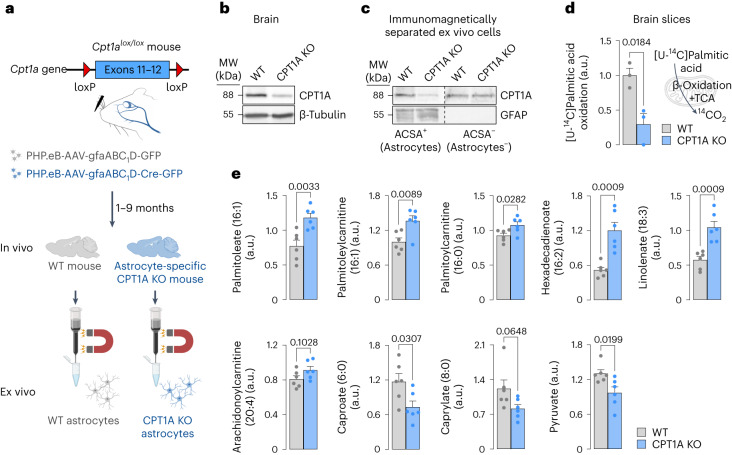
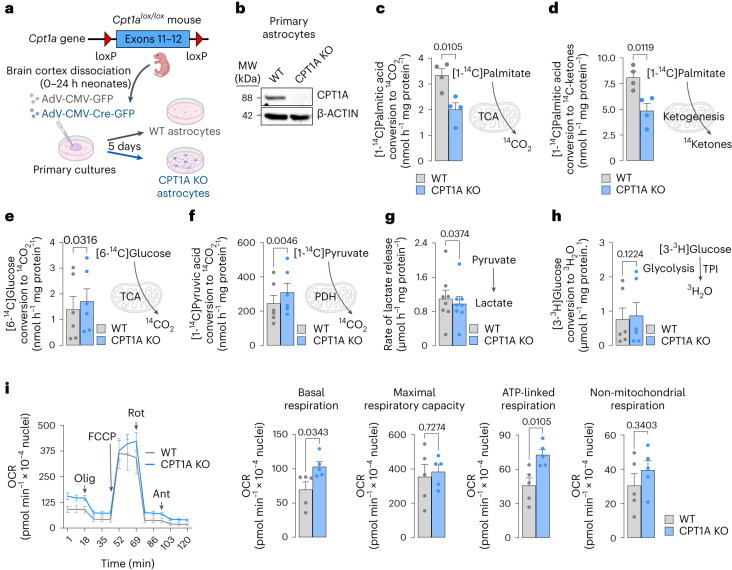
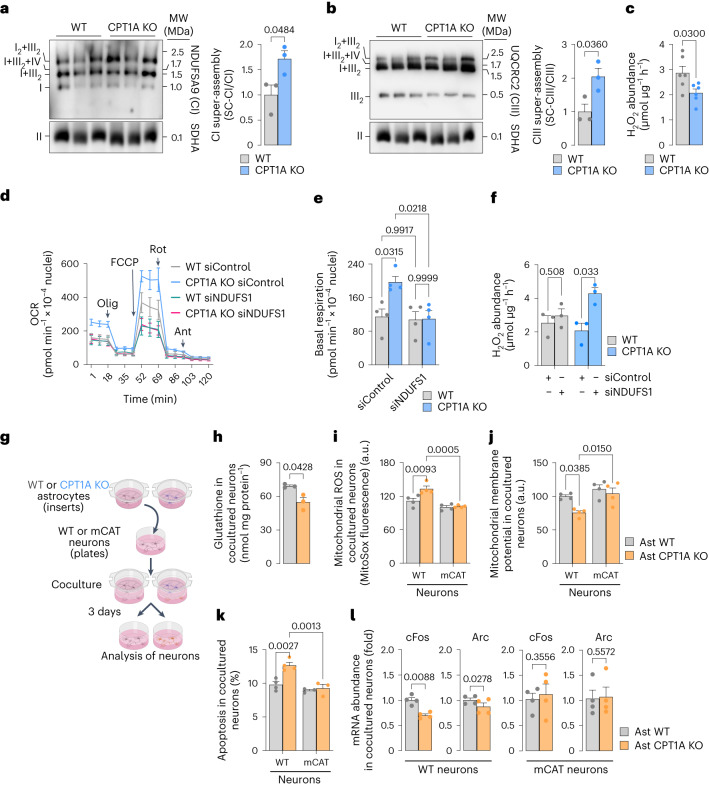
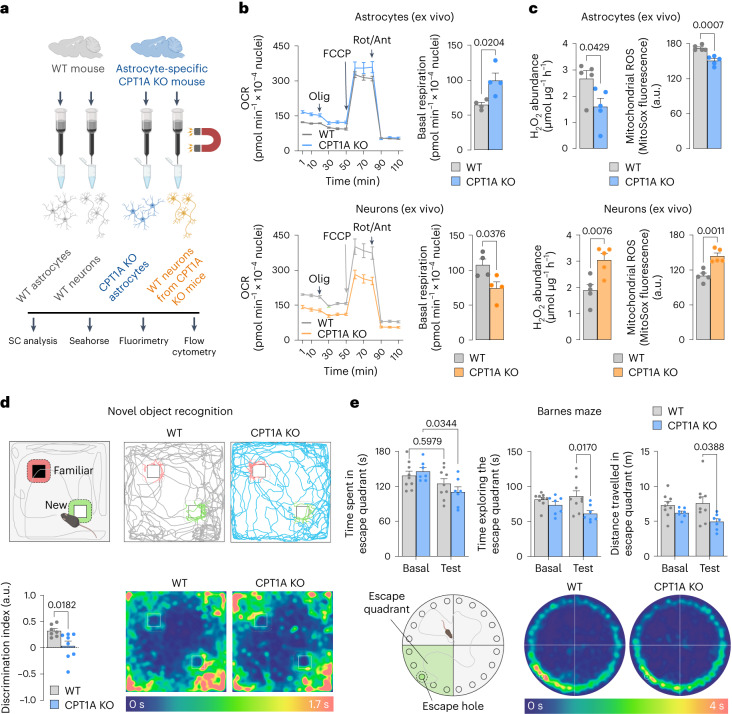
Having direct access to brain vasculature, astrocytes can take up available blood nutrients and metabolize them to fulfil their own energy needs and deliver metabolic intermediates to local synapses1,2. These glial cells should be, therefore, metabolically adaptable to swap different substrates. However, in vitro and in vivo studies consistently show that astrocytes are primarily glycolytic3-7, suggesting glucose is their main metabolic precursor. Notably, transcriptomic data8,9 and in vitro10 studies reveal that mouse astrocytes are capable of mitochondrially oxidizing fatty acids and that they can detoxify excess neuronal-derived fatty acids in disease models11,12. Still, the factual metabolic advantage of fatty acid use by astrocytes and its physiological impact on higher-order cerebral functions remain unknown. Here, we show that knockout of carnitine-palmitoyl transferase-1A (CPT1A)-a key enzyme of mitochondrial fatty acid oxidation-in adult mouse astrocytes causes cognitive impairment. Mechanistically, decreased fatty acid oxidation rewired astrocytic pyruvate metabolism to facilitate electron flux through a super-assembled mitochondrial respiratory chain, resulting in attenuation of reactive oxygen species formation. Thus, astrocytes naturally metabolize fatty acids to preserve the mitochondrial respiratory chain in an energetically inefficient disassembled conformation that secures signalling reactive oxygen species and sustains cognitive performance.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Kacem, K., Lacombe, P., Seylaz, J. & Bonvento, G. Structural organization of the perivascular astrocyte endfeet and their relationship with the endothelial glucose transporter: a confocal microscopy study. Glia23, 1–10 (1998). 10.1002/(SICI)1098-1136(199805)23:1<1::AID-GLIA1>3.0.CO;2-B - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
