

This is a preprint.
A critical role for cortical amygdala circuitry in shaping social encounters
- PMID: 37461537
- PMCID: PMC10350173
- DOI: 10.21203/rs.3.rs-3015820/v1
A critical role for cortical amygdala circuitry in shaping social encounters
Update in
-
A crucial role for the cortical amygdala in shaping social encounters.Nature. 2025 Mar;639(8056):1006-1015. doi: 10.1038/s41586-024-08540-4. Epub 2025 Feb 12. Nature. 2025. PMID: 39939764 Free PMC article.
Abstract
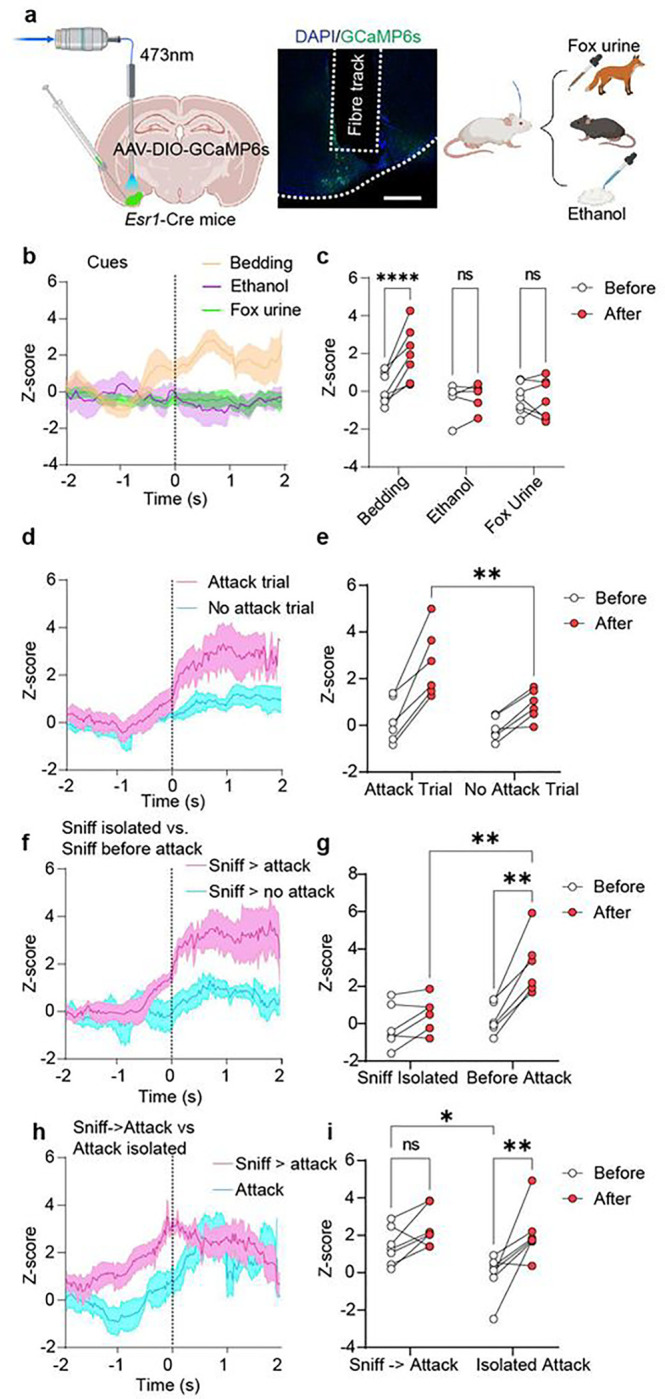
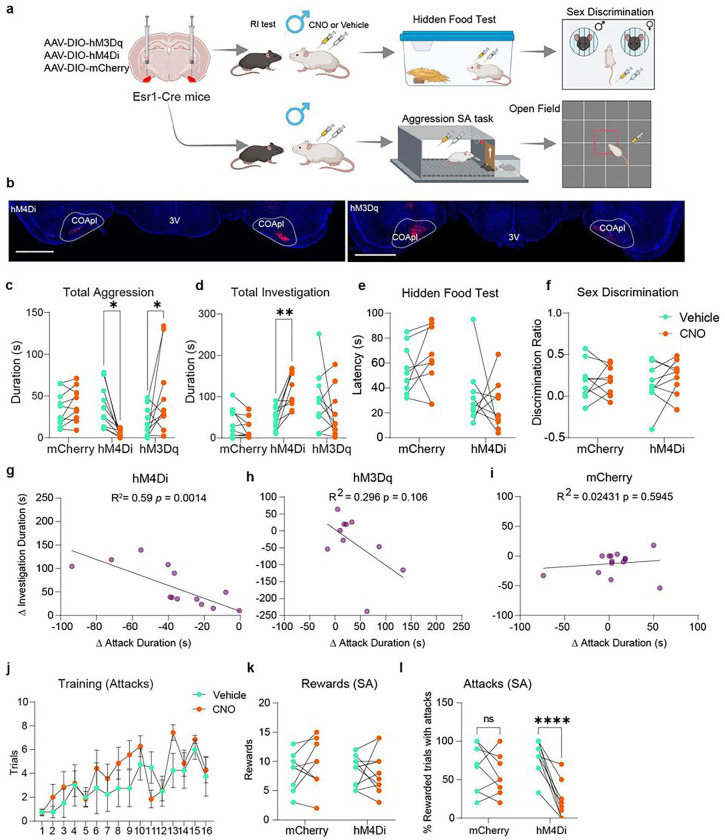
Aggression is an evolutionarily conserved behavior that controls social hierarchies and protects valuable resources like mates, food, and territory. In mice, aggressive behaviour can be broken down into an appetitive phase, which involves approach and investigation, and a consummatory phase, which involves biting, kicking, and wrestling. By performing an unsupervised weighted correlation network analysis on whole-brain c-Fos expression, we identified a cluster of brain regions including hypothalamic and amygdalar sub-regions and olfactory cortical regions highly co-activated in male, but not female aggressors (AGG). The posterolateral cortical amygdala (COApl), an extended olfactory structure, was found to be a hub region based on the number and strength of correlations with other regions in the cluster. Our data further show that estrogen receptor 1 (ESR1)-expressing cells in the COApl exhibit increased activity during attack behaviour, and during bouts of investigation which precede an attack, in male mice only. Chemogenetic or optogenetic inhibition of COApl ESR1 cells in AGG males reduces aggression and increases pro-social investigation without affecting social reward/reinforcement behavior. We further confirmed that COApl ESR1 projections to the ventrolateral portion of the ventromedial hypothalamus and central amygdala are necessary for these behaviours. Collectively, these data suggest that in aggressive males, COApl ESR1 cells respond specifically to social stimuli, thereby enhancing their salience and promoting attack behaviour.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
