

DeSUMOylation of chromatin-bound proteins limits the rapid transcriptional reprogramming induced by daunorubicin in acute myeloid leukemias
- PMID: 37462077
- PMCID: PMC10484680
- DOI: 10.1093/nar/gkad581
DeSUMOylation of chromatin-bound proteins limits the rapid transcriptional reprogramming induced by daunorubicin in acute myeloid leukemias
Abstract
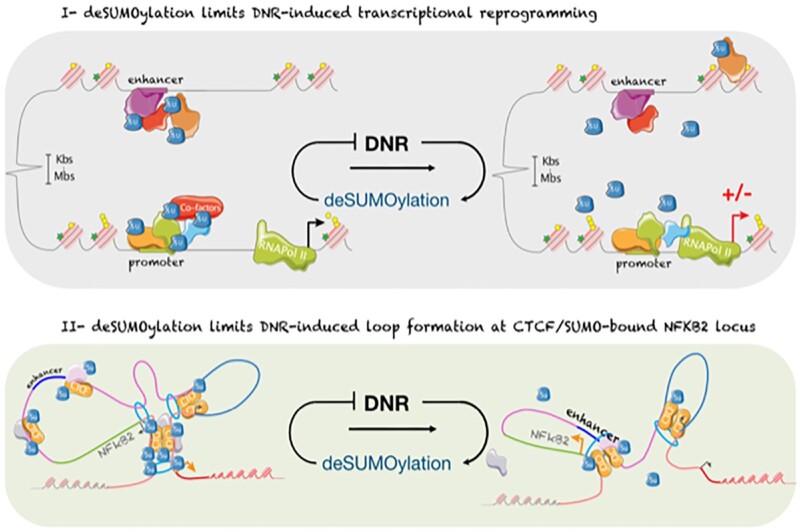
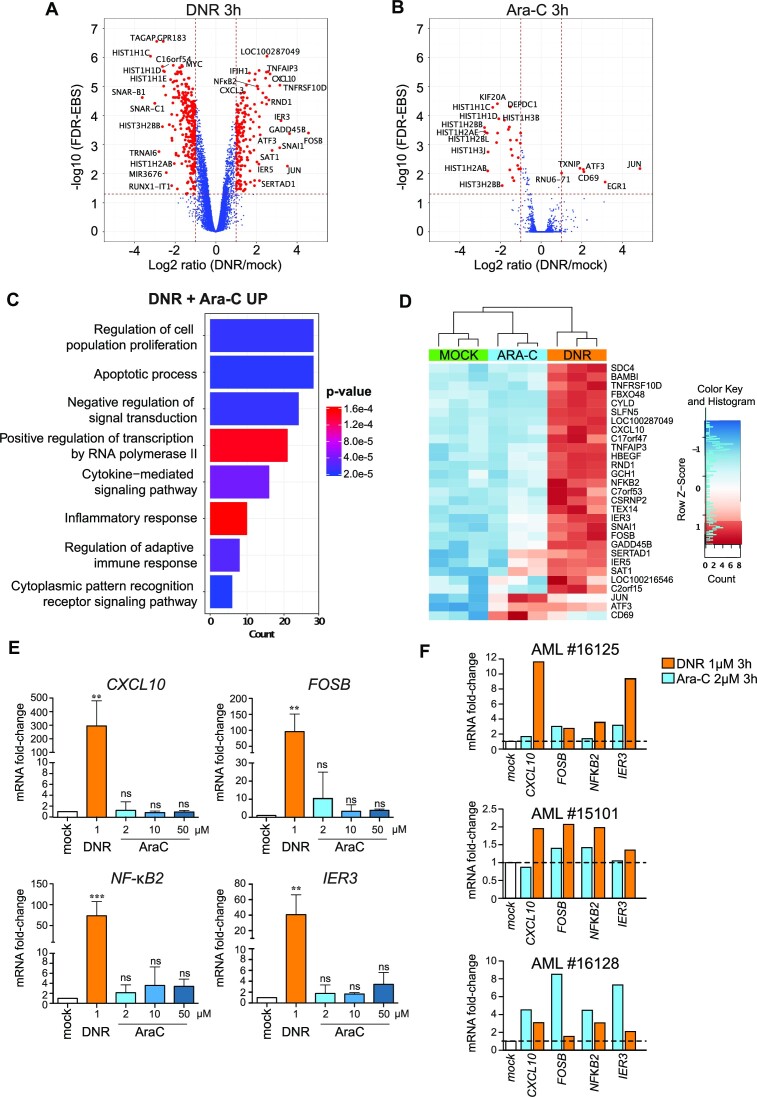
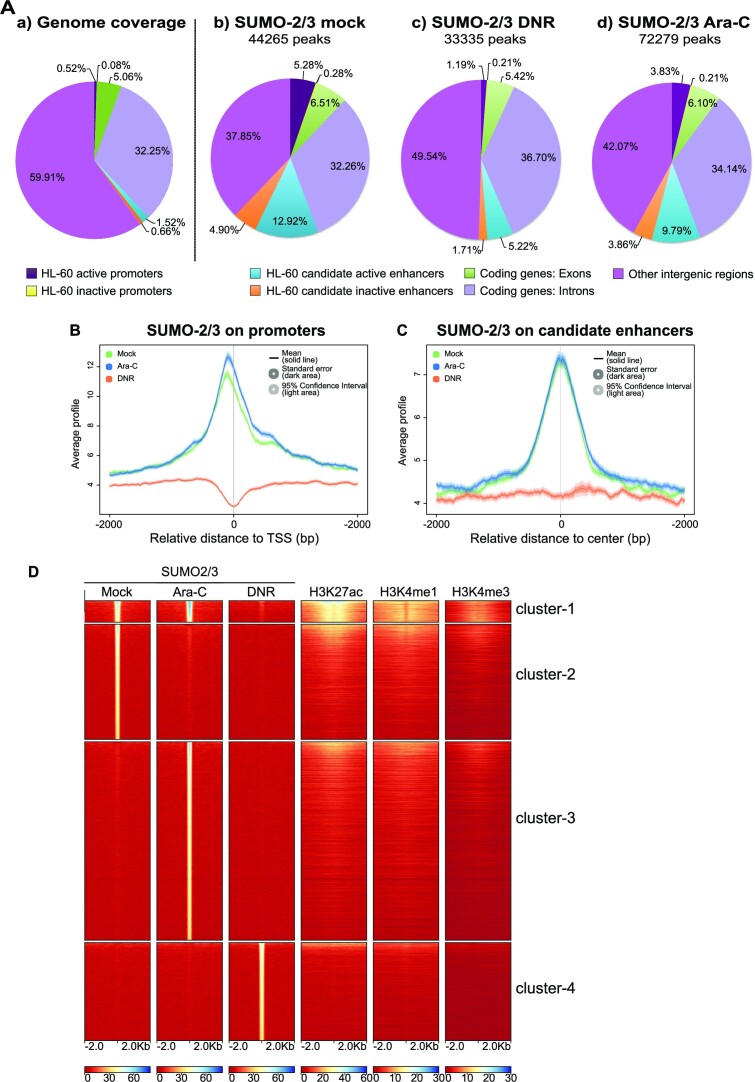
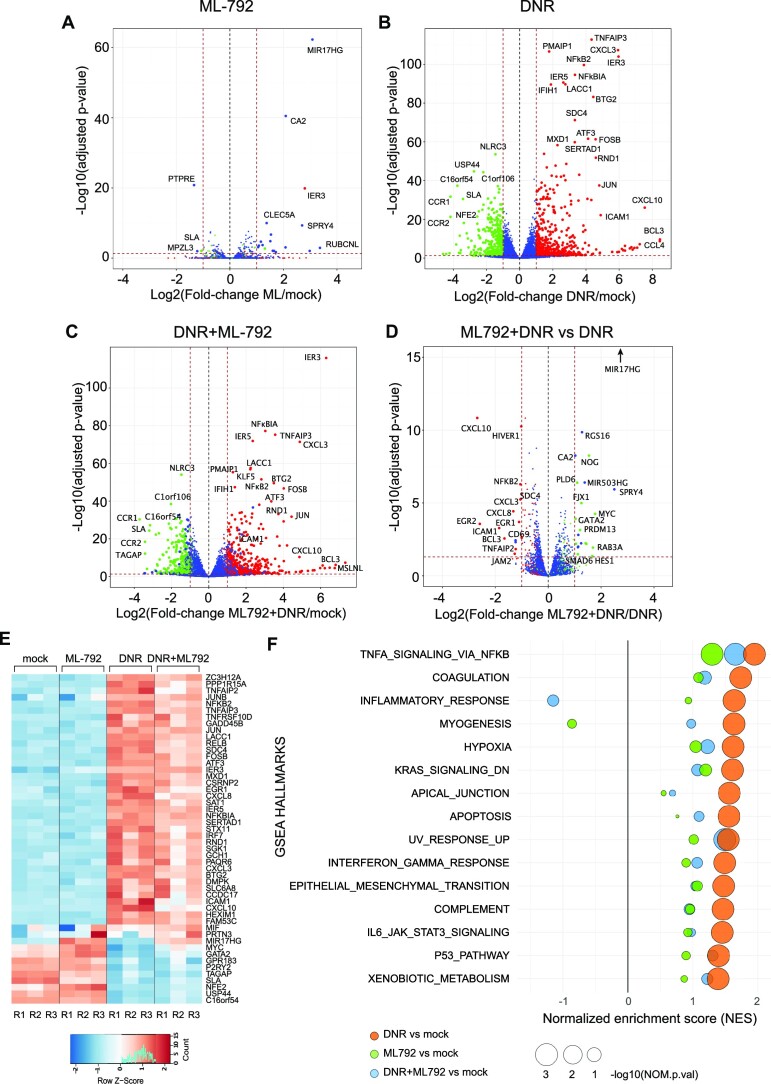
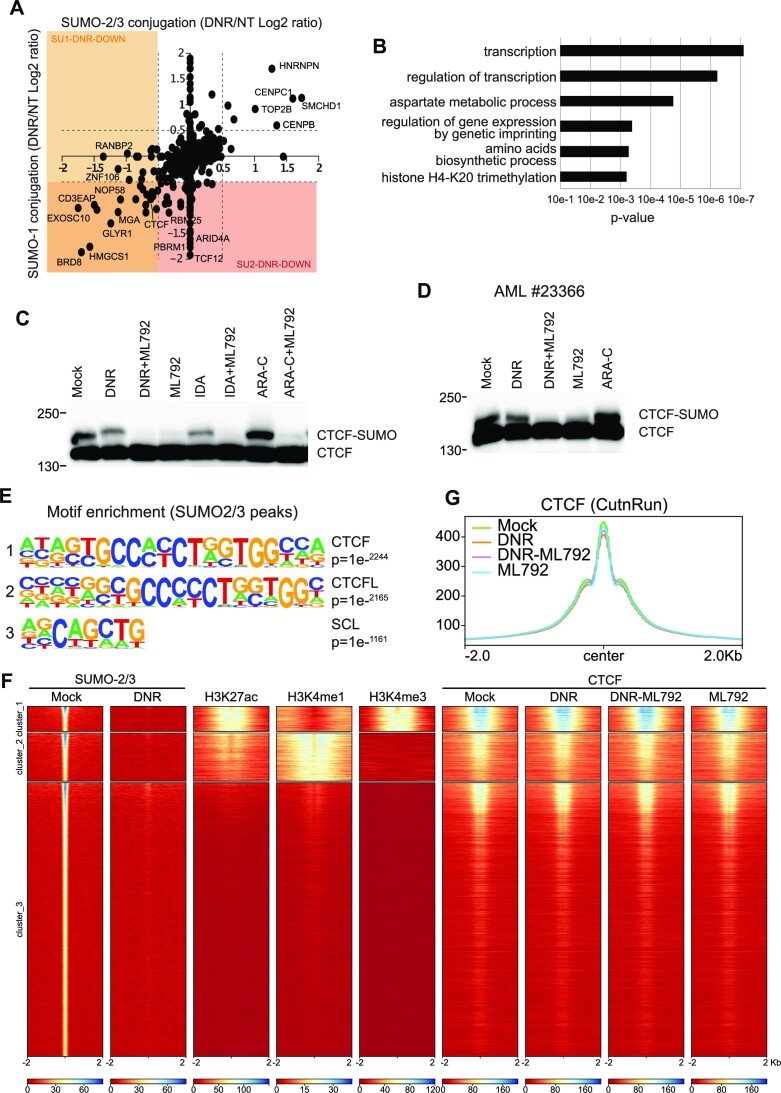
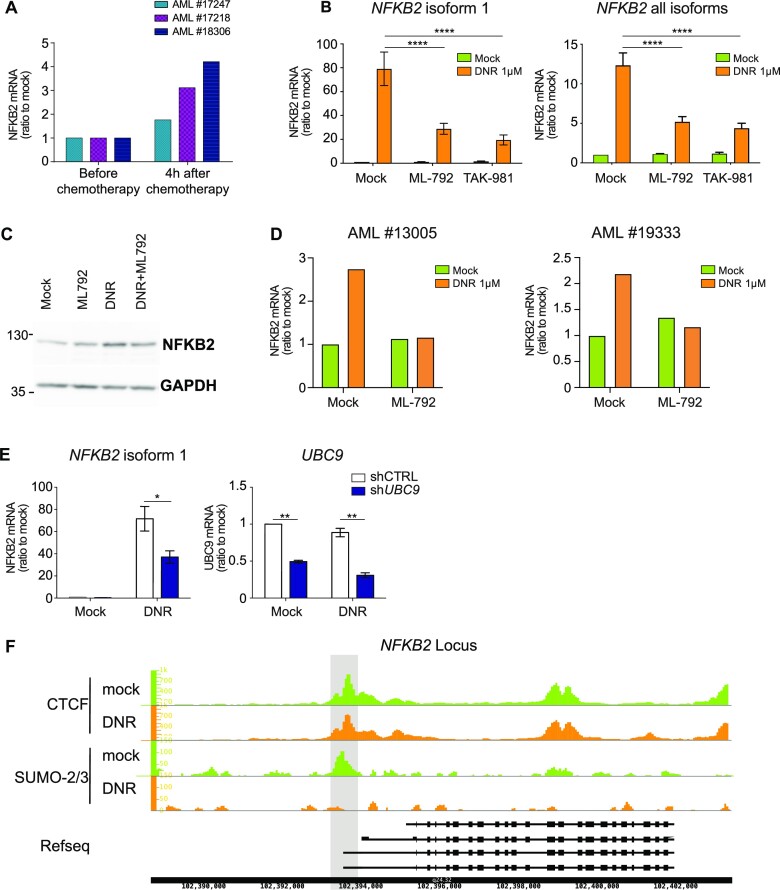
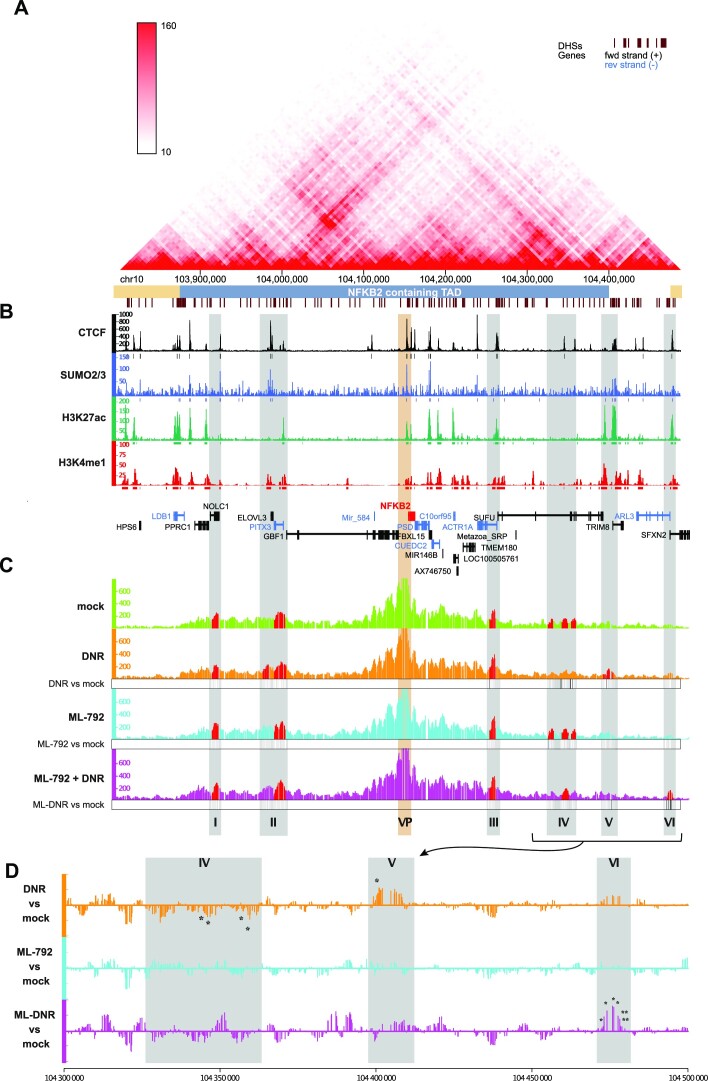
Genotoxicants have been used for decades as front-line therapies against cancer on the basis of their DNA-damaging actions. However, some of their non-DNA-damaging effects are also instrumental for killing dividing cells. We report here that the anthracycline Daunorubicin (DNR), one of the main drugs used to treat Acute Myeloid Leukemia (AML), induces rapid (3 h) and broad transcriptional changes in AML cells. The regulated genes are particularly enriched in genes controlling cell proliferation and death, as well as inflammation and immunity. These transcriptional changes are preceded by DNR-dependent deSUMOylation of chromatin proteins, in particular at active promoters and enhancers. Surprisingly, inhibition of SUMOylation with ML-792 (SUMO E1 inhibitor), dampens DNR-induced transcriptional reprogramming. Quantitative proteomics shows that the proteins deSUMOylated in response to DNR are mostly transcription factors, transcriptional co-regulators and chromatin organizers. Among them, the CCCTC-binding factor CTCF is highly enriched at SUMO-binding sites found in cis-regulatory regions. This is notably the case at the promoter of the DNR-induced NFKB2 gene. DNR leads to a reconfiguration of chromatin loops engaging CTCF- and SUMO-bound NFKB2 promoter with a distal cis-regulatory region and inhibition of SUMOylation with ML-792 prevents these changes.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Targeted inhibition of ULK1 enhances daunorubicin sensitivity in acute myeloid leukemia.Life Sci. 2020 Feb 15;243:117234. doi: 10.1016/j.lfs.2019.117234. Epub 2019 Dec 27. Life Sci. 2020. PMID: 31887299
-
NOXA-mediated degradation of MCL1 and BCL2L1 causes apoptosis of daunorubicin-treated human acute myeloid leukemia cells.J Cell Physiol. 2021 Nov;236(11):7356-7375. doi: 10.1002/jcp.30407. Epub 2021 May 13. J Cell Physiol. 2021. PMID: 33982799
-
Curcumin reduces expression of Bcl-2, leading to apoptosis in daunorubicin-insensitive CD34+ acute myeloid leukemia cell lines and primary sorted CD34+ acute myeloid leukemia cells.J Transl Med. 2011 May 19;9:71. doi: 10.1186/1479-5876-9-71. J Transl Med. 2011. PMID: 21595920 Free PMC article.
-
Equipotent doses of daunorubicin and idarubicin for AML: a meta-analysis of clinical trials versus in vitro estimation.Cancer Chemother Pharmacol. 2019 Jun;83(6):1105-1112. doi: 10.1007/s00280-019-03825-2. Epub 2019 Apr 9. Cancer Chemother Pharmacol. 2019. PMID: 30968179 Free PMC article. Review.
-
The use of liposomal daunorubicin (DaunoXome) in acute myeloid leukemia.Leuk Lymphoma. 2005 Jun;46(6):795-802. doi: 10.1080/10428190500052438. Leuk Lymphoma. 2005. PMID: 16019523 Review.
Cited by
-
Distinctive tumorigenic significance and innovative oncology targets of SUMOylation.Theranostics. 2024 May 19;14(8):3127-3149. doi: 10.7150/thno.97162. eCollection 2024. Theranostics. 2024. PMID: 38855173 Free PMC article. Review.
-
SUMOylation inhibitor TAK-981 (subasumstat) synergizes with 5-azacytidine in preclinical models of acute myeloid leukemia.Haematologica. 2024 Jan 1;109(1):98-114. doi: 10.3324/haematol.2023.282704. Haematologica. 2024. PMID: 37608777 Free PMC article.
-
Transient TKI-resistant CD44+pBAD+ blasts undergo intrinsic homeostatic adaptation to promote the survival of acute myeloid leukemia in vitro.Front Oncol. 2023 Nov 8;13:1286863. doi: 10.3389/fonc.2023.1286863. eCollection 2023. Front Oncol. 2023. PMID: 38023123 Free PMC article.
-
Targeting SUMOylation triggers interferon-ß-dependent activation of patient and allogenic Natural Killer cells in preclinical models of Acute Myeloid Leukemia.Mol Cancer Ther. 2025 Jul 15:10.1158/1535-7163.MCT-25-0504. doi: 10.1158/1535-7163.MCT-25-0504. Online ahead of print. Mol Cancer Ther. 2025. PMID: 40661051 Free PMC article.
-
The multifaceted nature of SUMOylation in heart disease and its therapeutic potential.Mol Cell Biochem. 2025 Aug;480(8):4725-4743. doi: 10.1007/s11010-025-05286-z. Epub 2025 Apr 27. Mol Cell Biochem. 2025. PMID: 40287894 Review.
References
-
- Khaled S., Al Malki M., Marcucci G.. Acute myeloid leukemia: biologic, prognostic, and therapeutic insights. Oncol. Williston Park N. 2016; 30:318–329. - PubMed
-
- Minotti G., Menna P., Salvatorelli E., Cairo G., Gianni L.. Anthracyclines: molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 2004; 56:185–229. - PubMed
-
- Grant S. Ara-C: cellular and molecular pharmacology. Adv. Cancer Res. 1998; 72:197–233. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
