

Dissecting the role of the NADPH oxidase NOX4 in TGF-beta signaling in hepatocellular carcinoma
- PMID: 37463530
- PMCID: PMC10372458
- DOI: 10.1016/j.redox.2023.102818
Dissecting the role of the NADPH oxidase NOX4 in TGF-beta signaling in hepatocellular carcinoma
Abstract
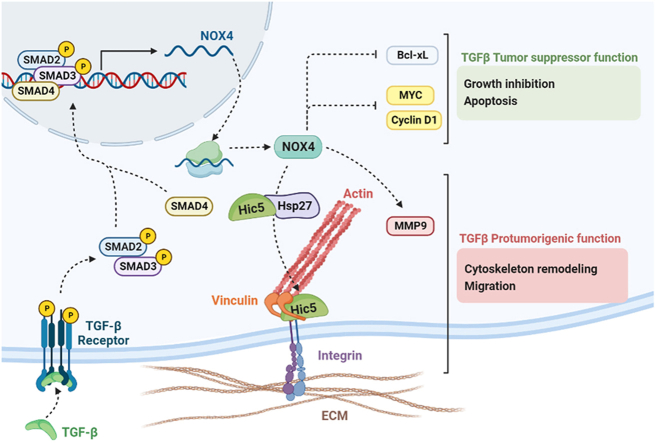
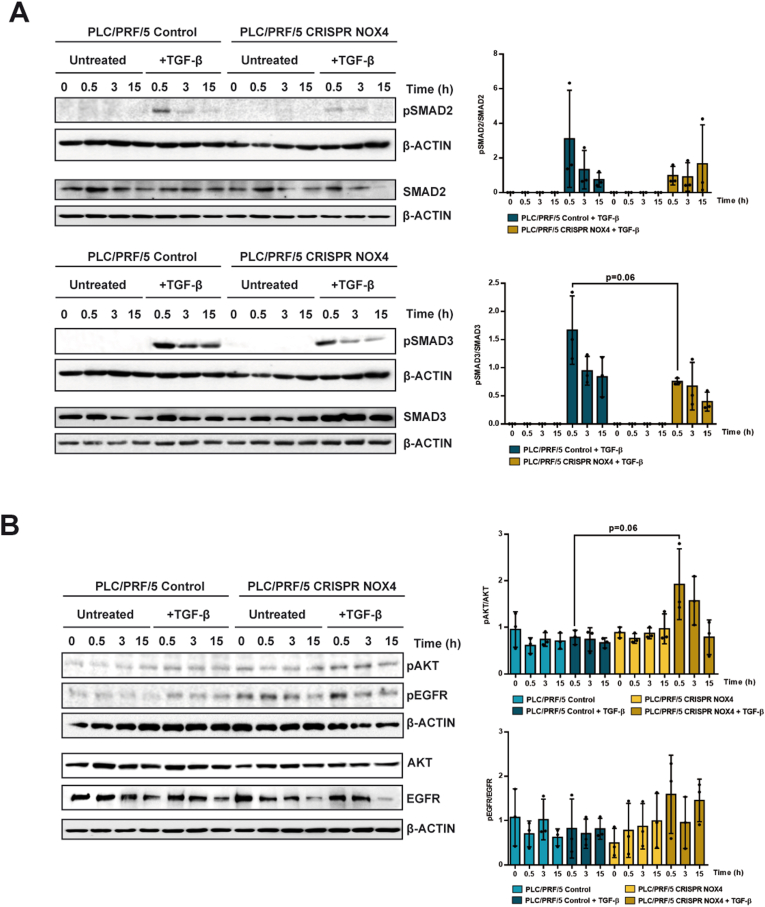
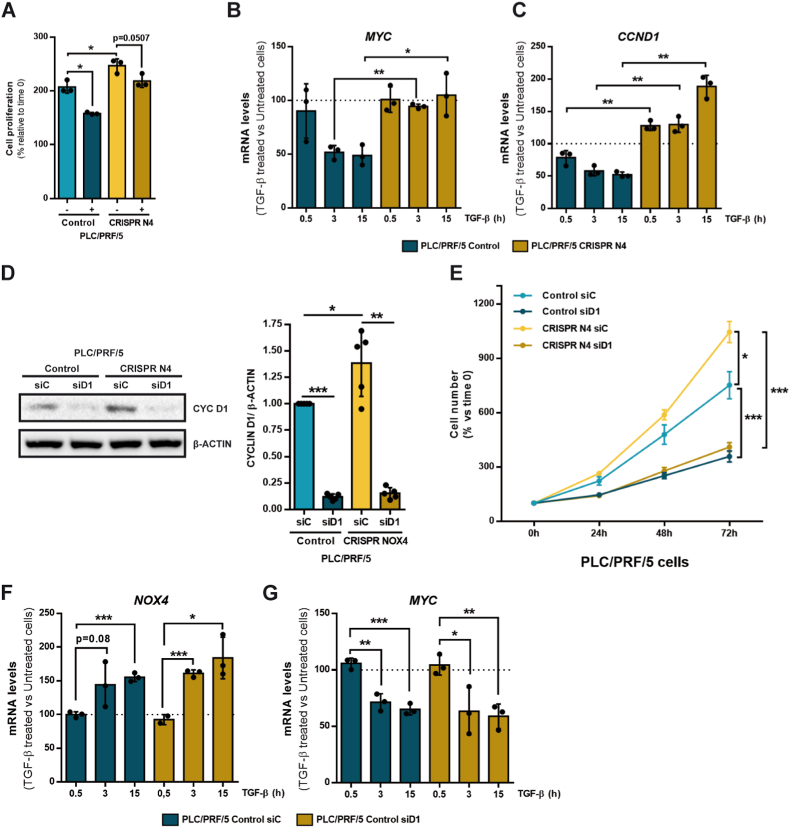
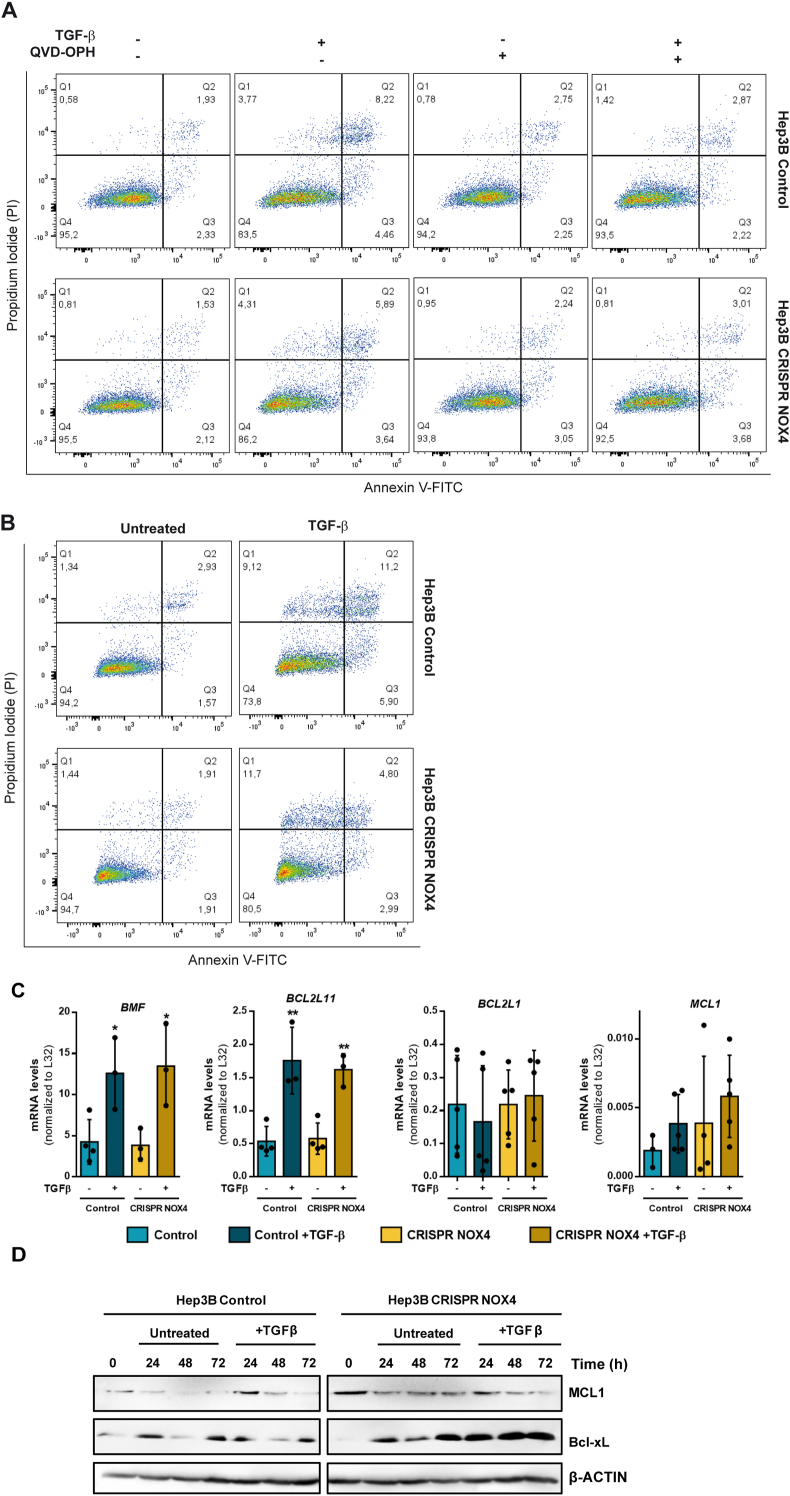
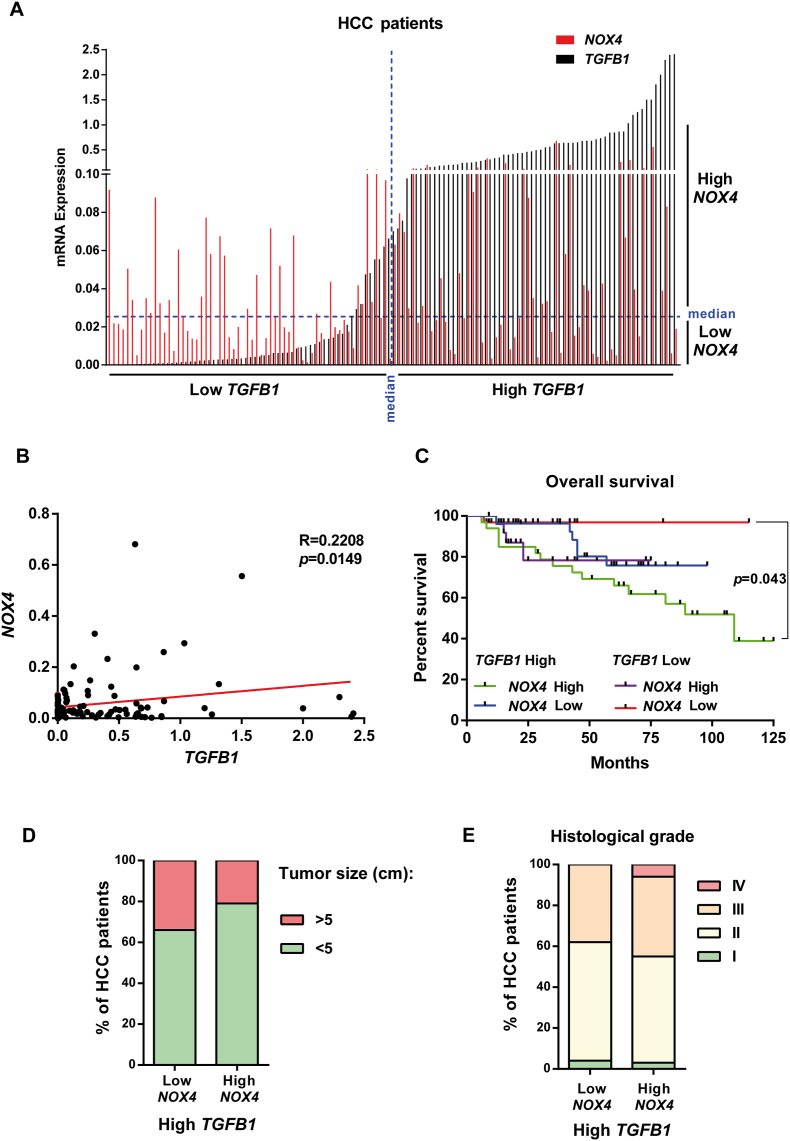
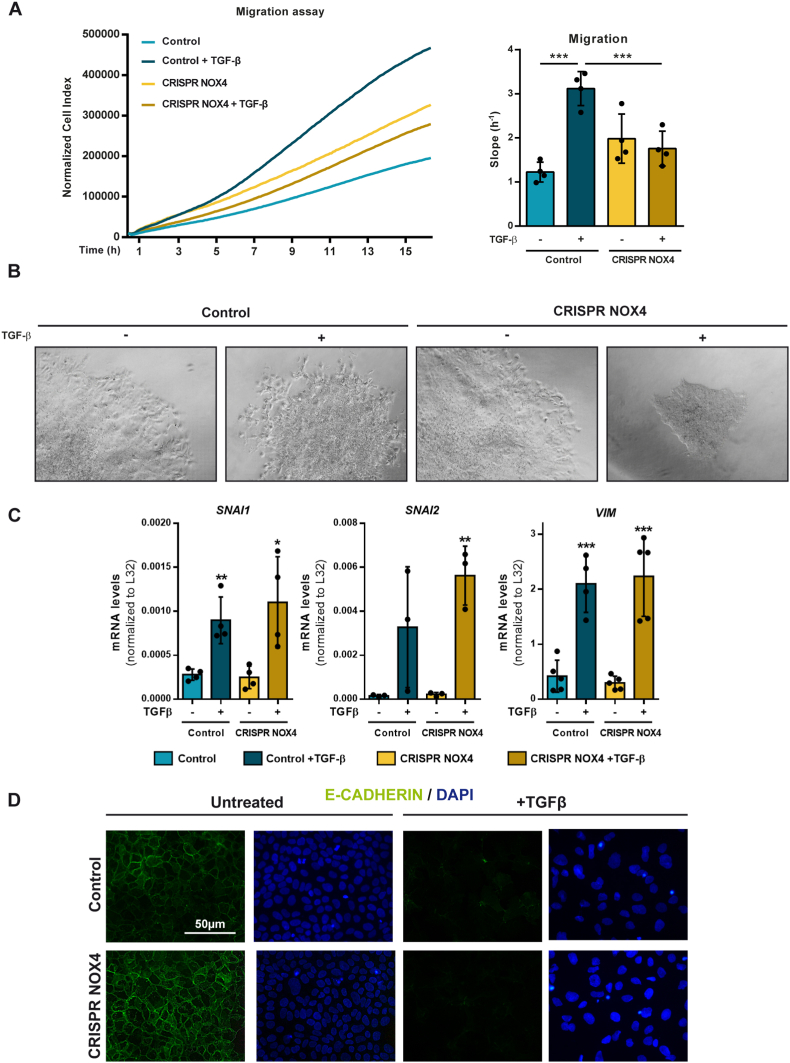
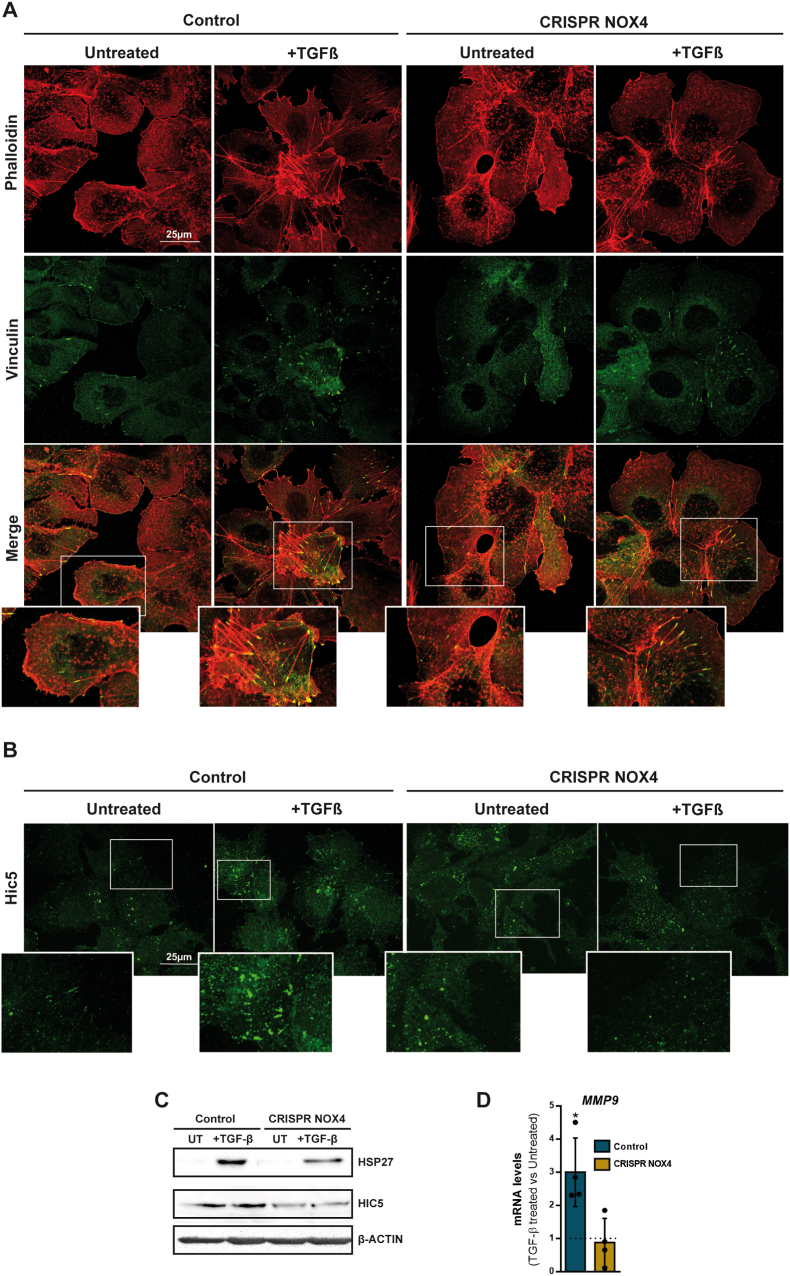
The NADPH oxidase NOX4 has been proposed as necessary for the apoptosis induced by the Transforming Growth Factor-beta (TGF-β) in hepatocytes and hepatocellular carcinoma (HCC) cells. However, whether NOX4 is required for TGF-β-induced canonical (SMADs) or non-canonical signals is not fully understood yet, neither its potential involvement in other parallel actions induced by TGF-β. In this work we have used CRISPR Cas9 technology to stable attenuate NOX4 expression in HCC cells. Results have indicated that NOX4 is required for an efficient SMAD2/3 phosphorylation in response to TGF-β, whereas non-canonical signals, such as the phosphorylation of the Epidermal Growth Receptor or AKT, are higher in NOX4 silenced cells. TGF-β-mediated inhibition of cell proliferation and viability is attenuated in NOX4 silenced cells, correlating with decreased response in terms of apoptosis, and maintenance of high expression of MYC and CYCLIN D1. These results would indicate that NOX4 is required for all the tumor suppressor actions of TGF-β in HCC. However, analysis in human HCC tumors has revealed a worse prognosis for patients showing high expression of TGF-β1-related genes concomitant with high expression of NOX4. Deepening into other tumorigenic actions of TGF-β that may contribute to tumor progression, we found that NOX4 is also required for TGF-β-induced migratory effects. The Epithelial-Mesenchymal transition (EMT) program does not appear to be affected by attenuation of NOX4 levels. However, TGF-β-mediated regulation of cytoskeleton dynamics and focal adhesions require NOX4, which is necessary for TGF-β-induced increase in the chaperone Hsp27 and correct subcellular localization of Hic-5 within focal adhesions, as well for upregulation of the metalloprotease MMP9. All these results together point to NOX4 as a key element in the whole TGF-β signaling in HCC cells, revealing an unknown role for NOX4 as tumor promoter in HCC patients presenting activation of the TGF-β pathway.
Keywords: HCC; Hepatocellular carcinoma; Liver cancer; NADPH oxidase; NOX4; TGF-Beta.
Copyright © 2023 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Isabel Fabregat reports financial support was provided by Agencia Estatal de Investigación, Ministry of Science and Innovation, Spain. Rut Espinosa-Sotelo reports financial support was provided by Agencia Estatal de Investigación, Ministry of Science and Innovation, Spain. Isabel Fabregat reports financial support was provided by Asociación Española contra el Cáncer (AECC), Spain. Isabel Fabregat reports financial support was provided by Agència de Gestió d’Ajuts Universitaris i de Recerca (AGAUR, Generalitat de Catalunya). Isabel Fabregat reports financial support was provided by CIBER, National Biomedical Research Institute, Instituto de Salud Carlos III, Spain.
Figures
References
-
- Carmona-Cuenca I., Roncero C., Sancho P., Caja L., Fausto N., Fernández M., et al. Upregulation of the NADPH oxidase NOX4 by TGF-beta in hepatocytes is required for its pro-apoptotic activity. Journal of Hepatology. diciembre de. 2008;49(6):965–976. - PubMed
-
- Caja L., Sancho P., Bertran E., Iglesias-Serret D., Gil J., Fabregat I. Overactivation of the MEK/ERK pathway in liver tumor cells confers resistance to TGF-β-induced cell death through impairing up-regulation of the NADPH oxidase NOX4. Cancer Research. 1 de octubre de. 2009;69(19):7595–7602. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
