

Salicylic acid metabolism and signalling coordinate senescence initiation in aspen in nature
- PMID: 37463905
- PMCID: PMC10354028
- DOI: 10.1038/s41467-023-39564-5
Salicylic acid metabolism and signalling coordinate senescence initiation in aspen in nature
Abstract
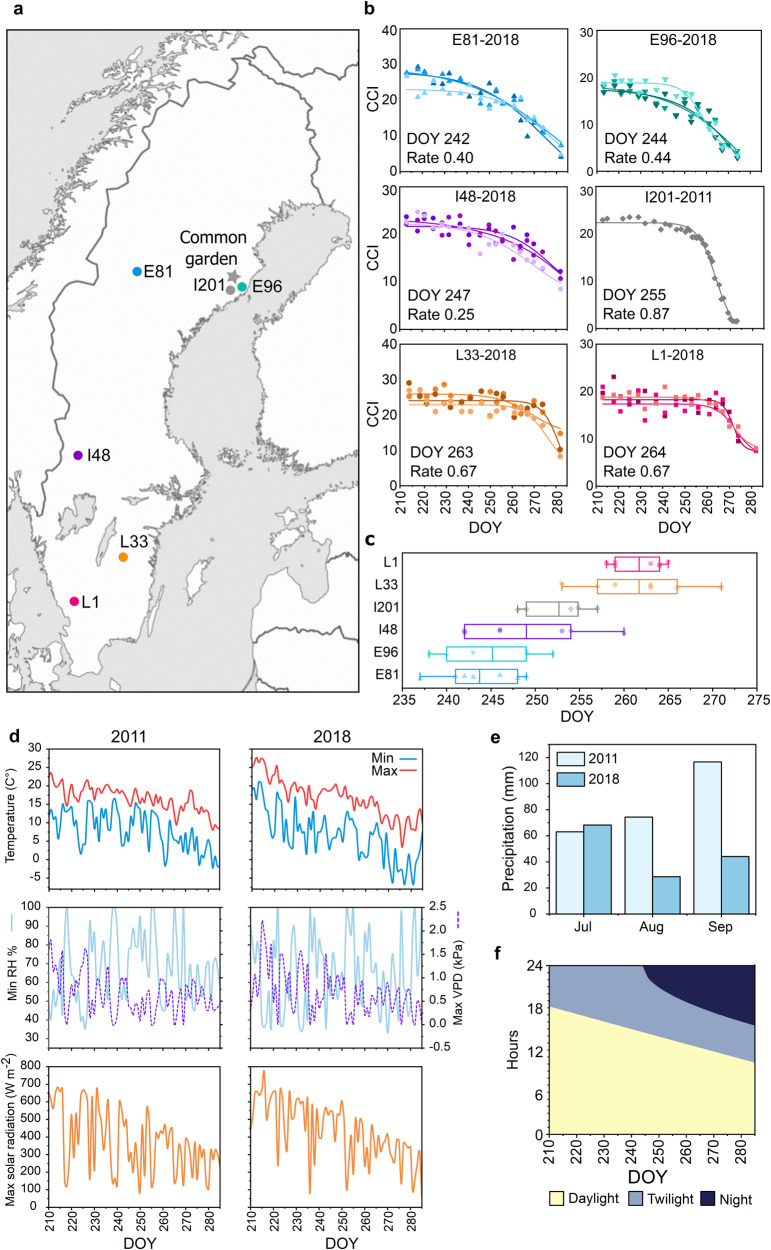
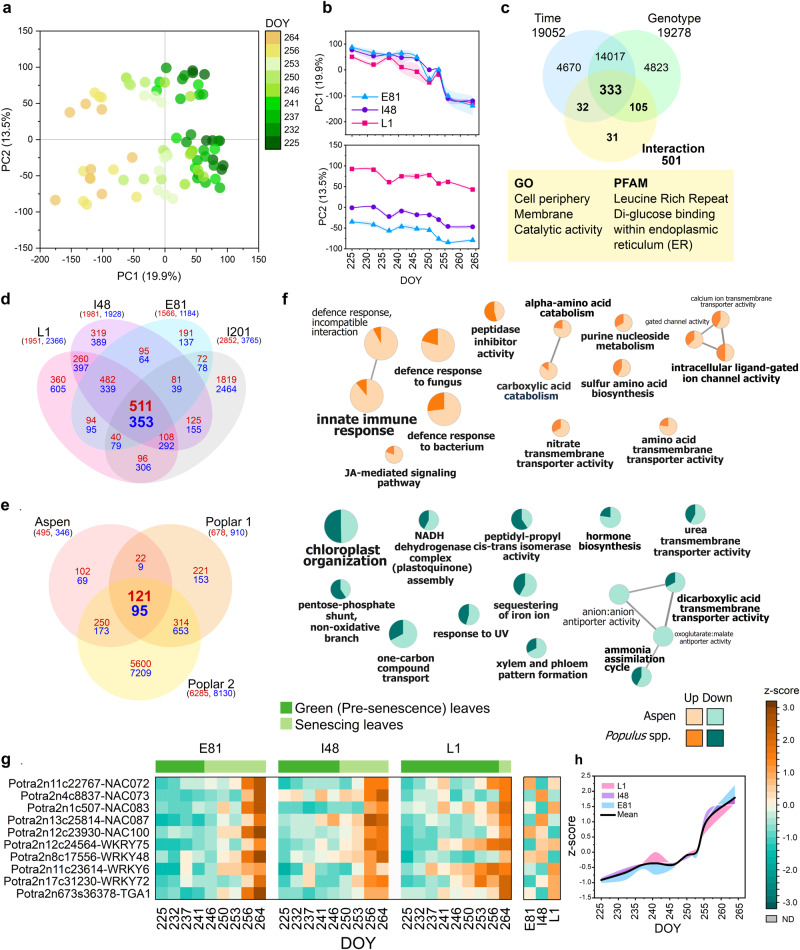
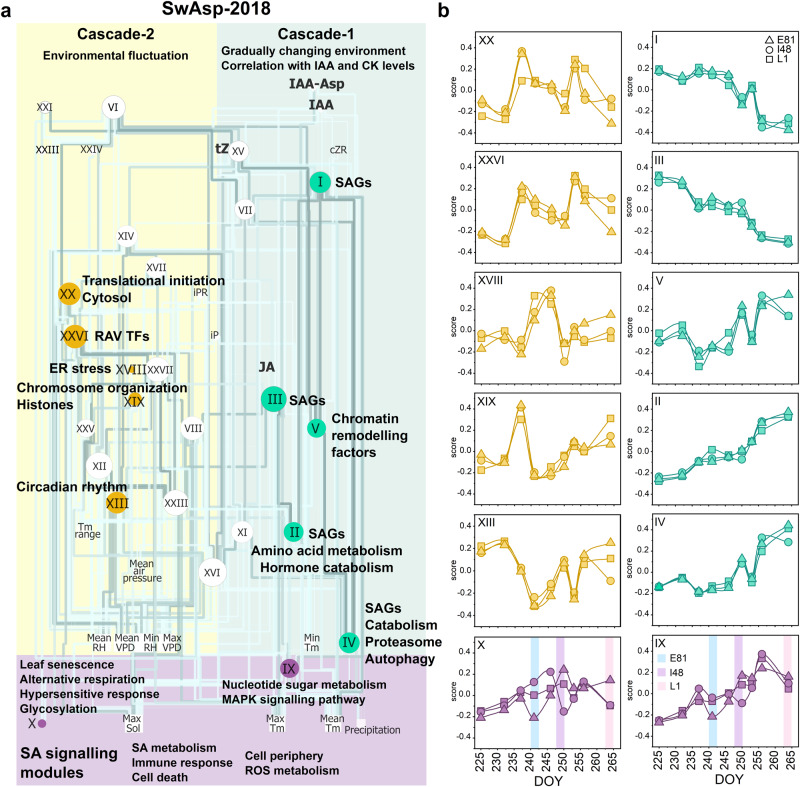
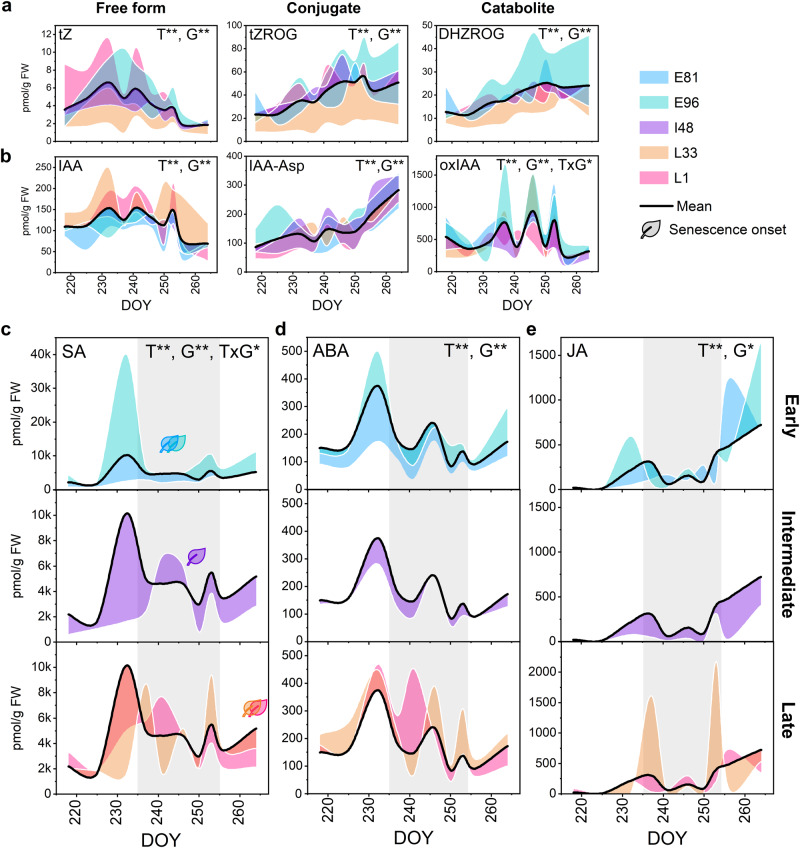
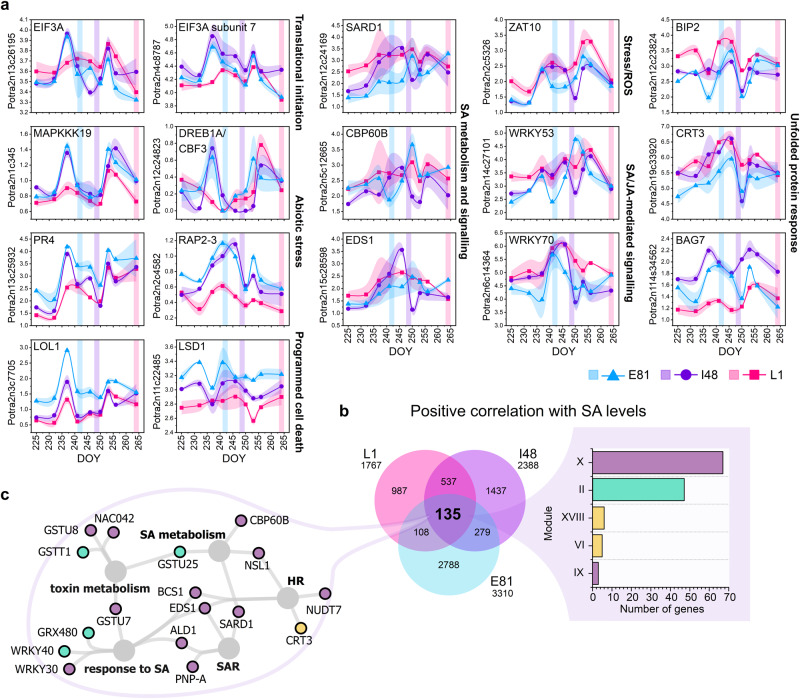
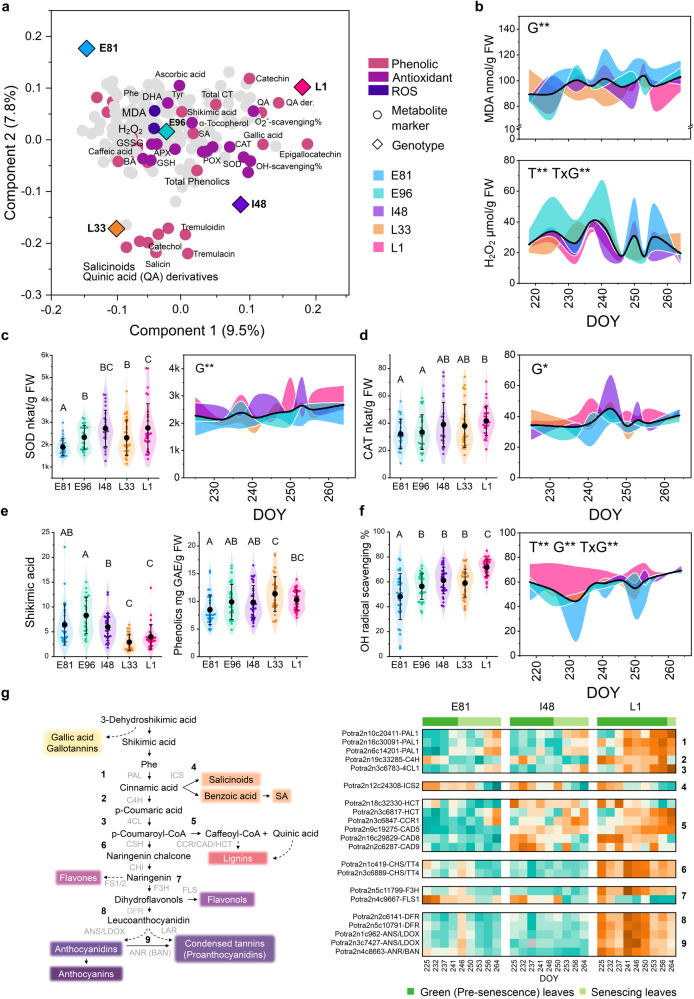
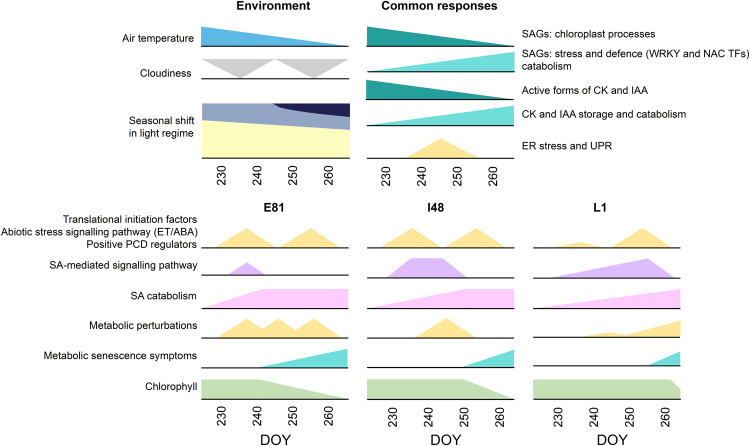
Deciduous trees exhibit a spectacular phenomenon of autumn senescence driven by the seasonality of their growth environment, yet there is no consensus which external or internal cues trigger it. Senescence starts at different times in European aspen (Populus tremula L.) genotypes grown in same location. By integrating omics studies, we demonstrate that aspen genotypes utilize similar transcriptional cascades and metabolic cues to initiate senescence, but at different times during autumn. The timing of autumn senescence initiation appeared to be controlled by two consecutive "switches"; 1) first the environmental variation induced the rewiring of the transcriptional network, stress signalling pathways and metabolic perturbations and 2) the start of senescence process was defined by the ability of the genotype to activate and sustain stress tolerance mechanisms mediated by salicylic acid. We propose that salicylic acid represses the onset of leaf senescence in stressful natural conditions, rather than promoting it as often observed in annual plants.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
The control of autumn senescence in European aspen.Plant Physiol. 2009 Apr;149(4):1982-91. doi: 10.1104/pp.108.133249. Epub 2009 Feb 6. Plant Physiol. 2009. PMID: 19201914 Free PMC article.
-
Autumn senescence in aspen is not triggered by day length.Physiol Plant. 2018 Jan;162(1):123-134. doi: 10.1111/ppl.12593. Epub 2017 Jul 11. Physiol Plant. 2018. PMID: 28591431
-
Stem girdling affects the onset of autumn senescence in aspen in interaction with metabolic signals.Physiol Plant. 2021 May;172(1):201-217. doi: 10.1111/ppl.13319. Epub 2021 Jan 7. Physiol Plant. 2021. PMID: 33368469 Free PMC article.
-
Signal transduction in leaf senescence.Plant Mol Biol. 2013 Aug;82(6):539-45. doi: 10.1007/s11103-012-9980-4. Epub 2012 Oct 25. Plant Mol Biol. 2013. PMID: 23096425 Review.
-
Hormonal regulation of leaf senescence through integration of developmental and stress signals.Plant Mol Biol. 2013 Aug;82(6):547-61. doi: 10.1007/s11103-013-0043-2. Epub 2013 Mar 16. Plant Mol Biol. 2013. PMID: 23504405 Review.
Cited by
-
Advances in Roles of Salicylic Acid in Plant Tolerance Responses to Biotic and Abiotic Stresses.Plants (Basel). 2023 Oct 4;12(19):3475. doi: 10.3390/plants12193475. Plants (Basel). 2023. PMID: 37836215 Free PMC article. Review.
-
CGD: a multi-omics database for Chrysanthemum genomic and biological research.Hortic Res. 2024 Aug 21;11(11):uhae238. doi: 10.1093/hr/uhae238. eCollection 2024 Nov. Hortic Res. 2024. PMID: 39512782 Free PMC article.
-
In vivo dynamics of indole- and phenol-derived plant hormones: Long-term, continuous, and minimally invasive phytohormone sensor.Sci Adv. 2025 Apr 18;11(16):eads8733. doi: 10.1126/sciadv.ads8733. Epub 2025 Apr 18. Sci Adv. 2025. PMID: 40249809 Free PMC article.
-
Simulation study of factors affecting the accuracy of transcriptome models under complex environments.Plant Mol Biol. 2025 Mar 28;115(2):52. doi: 10.1007/s11103-025-01578-6. Plant Mol Biol. 2025. PMID: 40153098
-
Salicylic acid accumulation: emerging molecular players and novel perspectives on plant development and nutrition.J Exp Bot. 2025 May 10;76(7):1950-1969. doi: 10.1093/jxb/erae309. J Exp Bot. 2025. PMID: 39028261 Free PMC article. Review.
References
-
- Woo HR, Kim HJ, Nam HG, Lim PO. Plant leaf senescence and death – regulation by multiple layers of control and implications for aging in general. J. Cell Sci. 2013;126:4823–4833. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
