

Cancer cell employs a microenvironmental neural signal trans-activating nucleus-mitochondria coordination to acquire stemness
- PMID: 37463926
- PMCID: PMC10354099
- DOI: 10.1038/s41392-023-01487-4
Cancer cell employs a microenvironmental neural signal trans-activating nucleus-mitochondria coordination to acquire stemness
Erratum in
-
Correction: Cancer cell employs a microenvironmental neural signal trans-activating nucleus-mitochondria coordination to acquire stemness.Signal Transduct Target Ther. 2025 Sep 2;10(1):289. doi: 10.1038/s41392-025-02389-3. Signal Transduct Target Ther. 2025. PMID: 40897726 Free PMC article. No abstract available.
Abstract
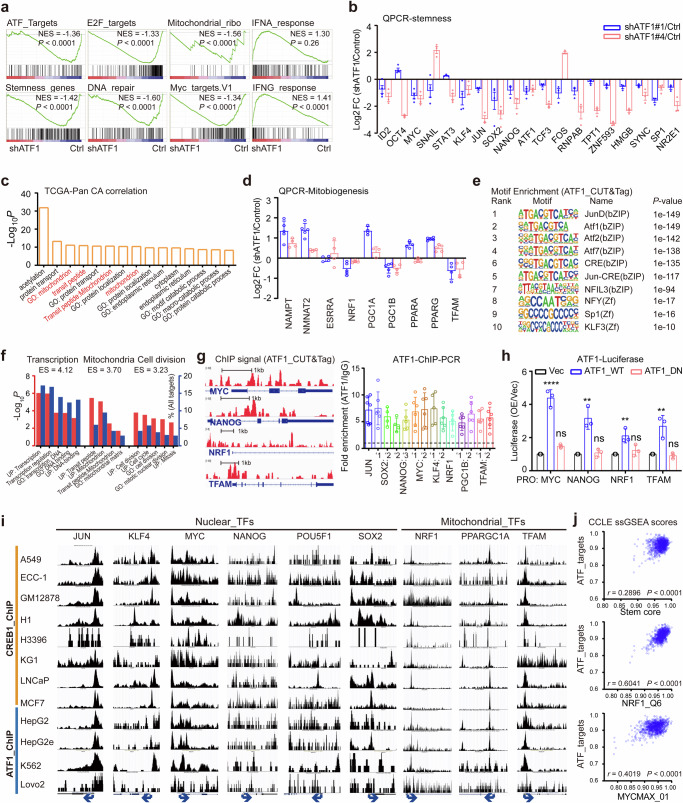
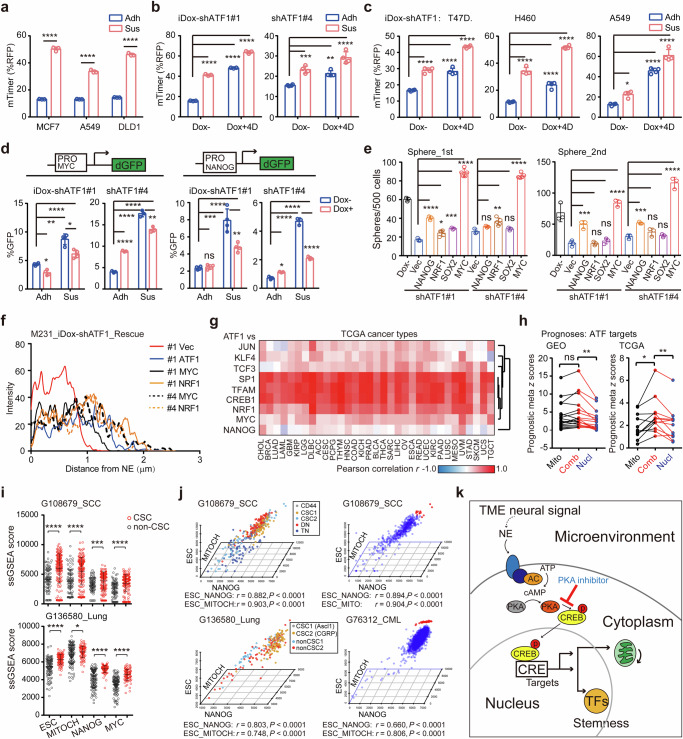
Cancer cell receives extracellular signal inputs to obtain a stem-like status, yet how tumor microenvironmental (TME) neural signals steer cancer stemness to establish the hierarchical tumor architectures remains elusive. Here, a pan-cancer transcriptomic screening for 10852 samples of 33 TCGA cancer types reveals that cAMP-responsive element (CRE) transcription factors are convergent activators for cancer stemness. Deconvolution of transcriptomic profiles, specification of neural markers and illustration of norepinephrine dynamics uncover a bond between TME neural signals and cancer-cell CRE activity. Specifically, neural signal norepinephrine potentiates the stemness of proximal cancer cells by activating cAMP-CRE axis, where ATF1 serves as a conserved hub. Upon activation by norepinephrine, ATF1 potentiates cancer stemness by coordinated trans-activation of both nuclear pluripotency factors MYC/NANOG and mitochondrial biogenesis regulators NRF1/TFAM, thereby orchestrating nuclear reprograming and mitochondrial rejuvenating. Accordingly, single-cell transcriptomes confirm the coordinated activation of nuclear pluripotency with mitochondrial biogenesis in cancer stem-like cells. These findings elucidate that cancer cell acquires stemness via a norepinephrine-ATF1 driven nucleus-mitochondria collaborated program, suggesting a spatialized stemness acquisition by hijacking microenvironmental neural signals.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
