

Identification of genes associated with the regulation of cold tolerance and the RNA movement in the grafted apple
- PMID: 37463950
- PMCID: PMC10354032
- DOI: 10.1038/s41598-023-38571-2
Identification of genes associated with the regulation of cold tolerance and the RNA movement in the grafted apple
Abstract
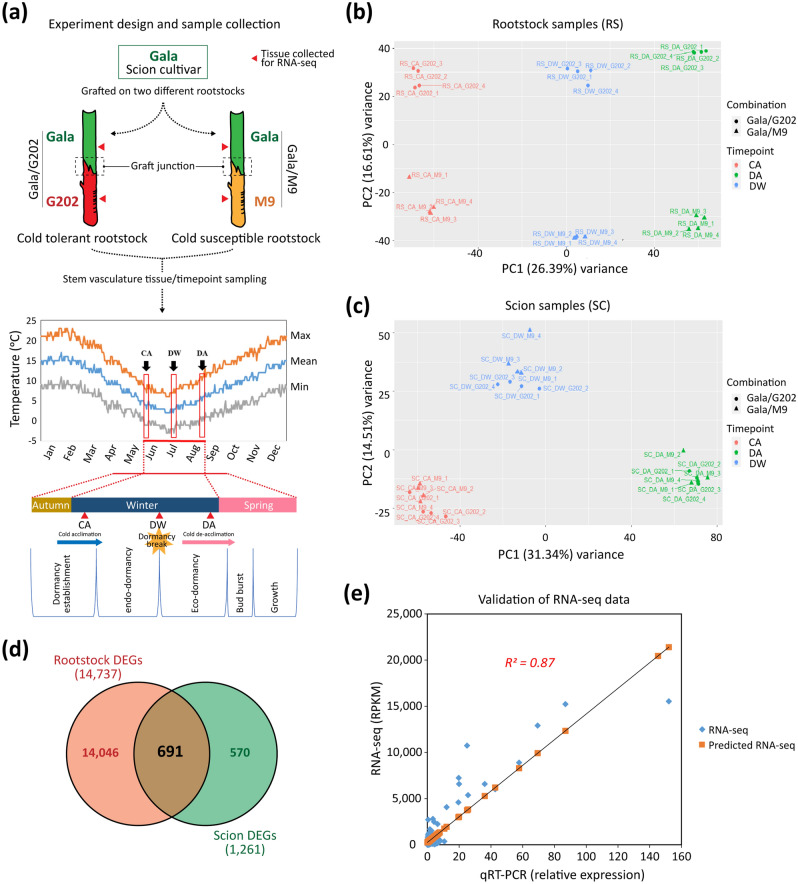
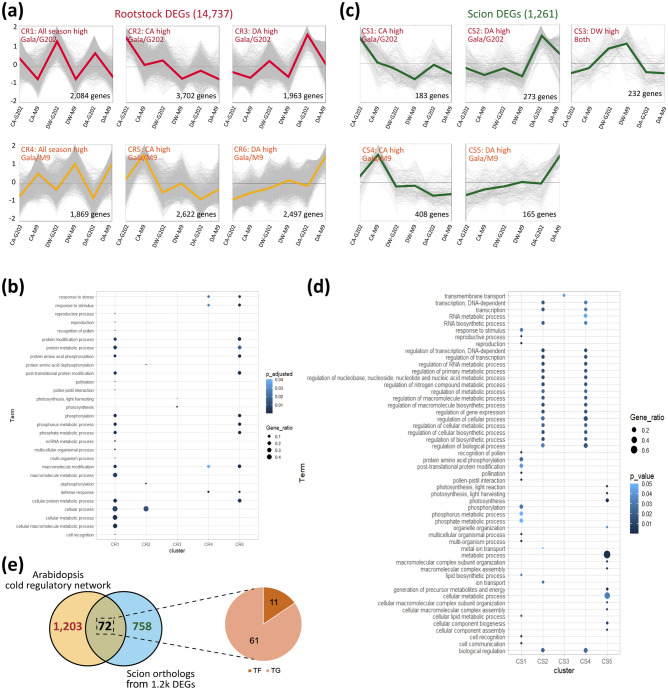
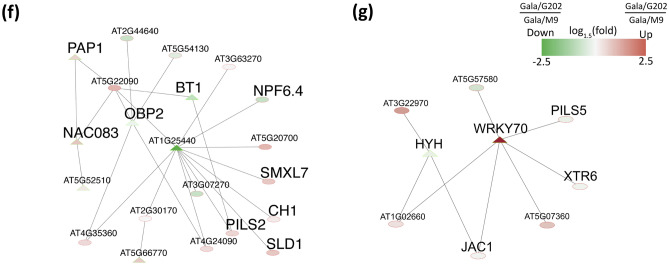
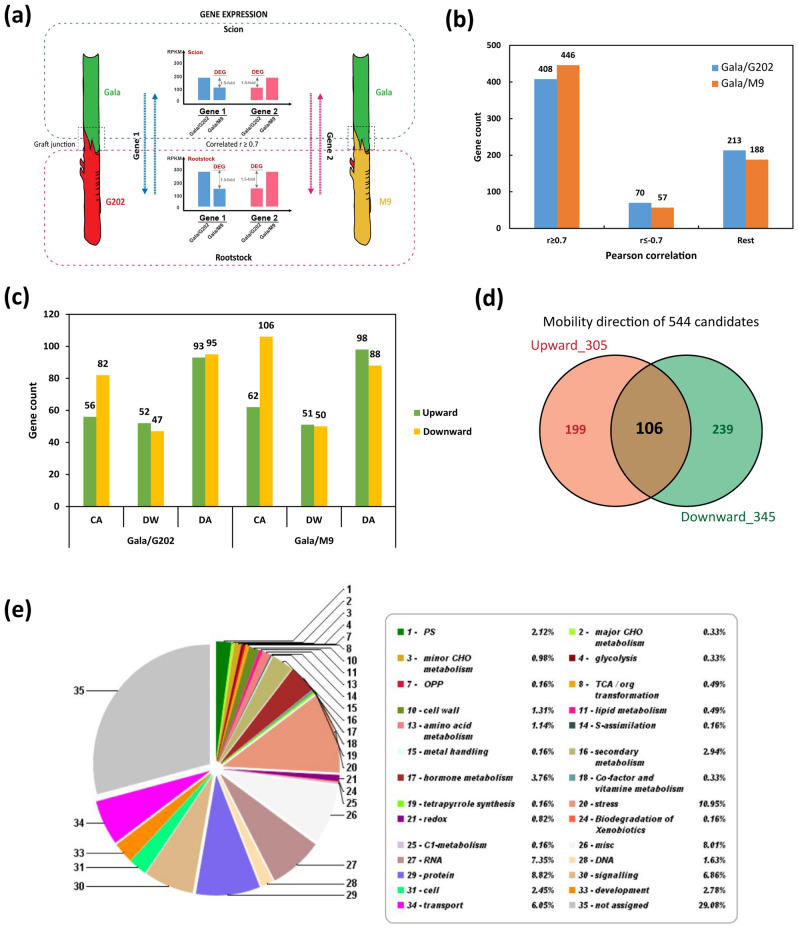
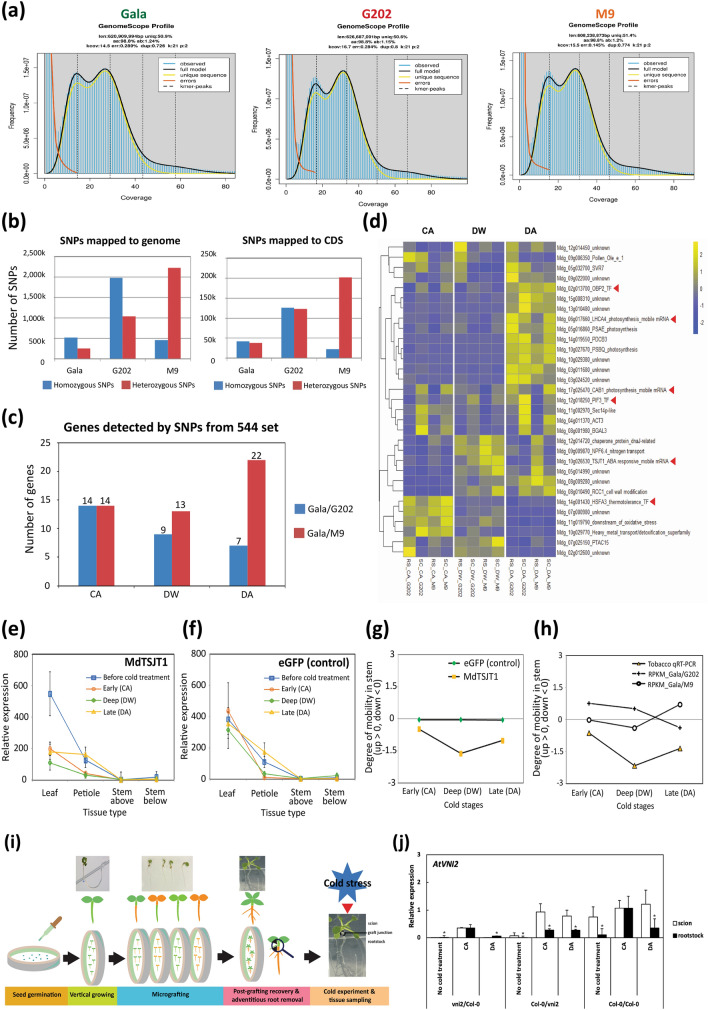
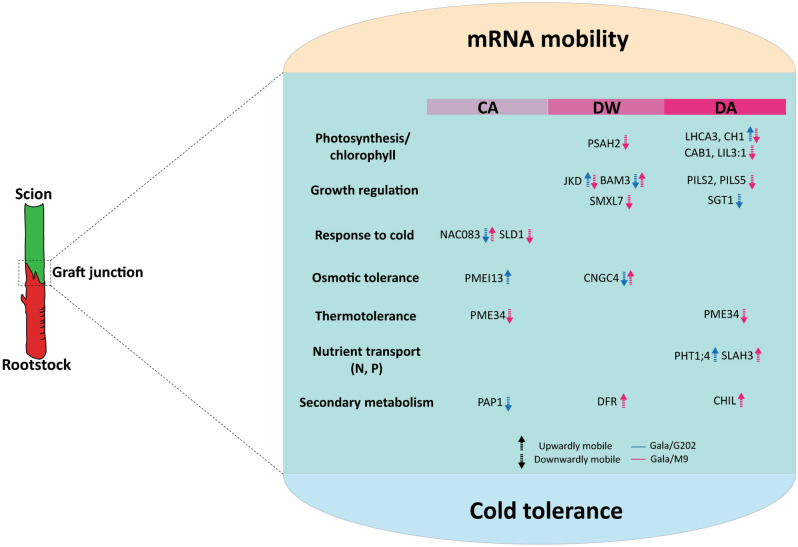
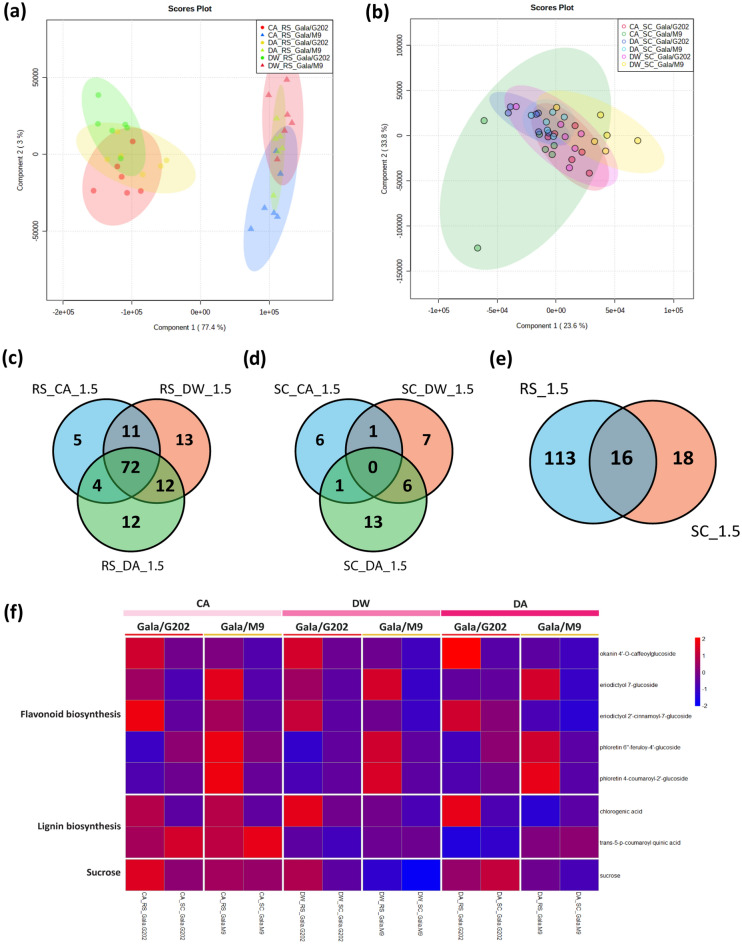
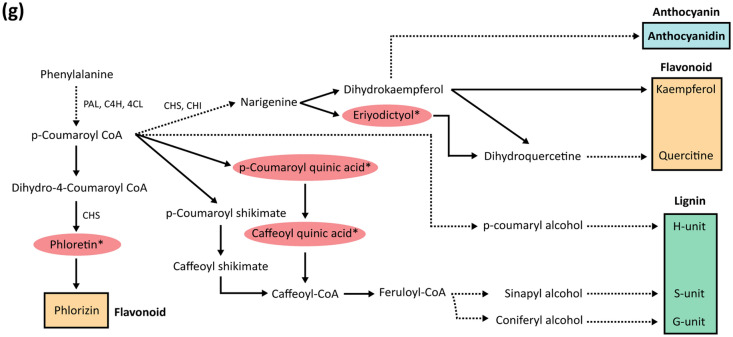
In grafted apple, rootstock-derived signals influence scion cold tolerance by initiating physiological changes to survive over the winter. To understand the underlying molecular interactions between scion and rootstock responsive to cold, we developed transcriptomics and metabolomics data in the stems of two scion/rootstock combinations, 'Gala'/'G202' (cold resistant rootstock) and 'Gala'/'M9' (cold susceptible rootstock). Outer layers of scion and rootstock stem, including vascular tissues, were collected from the field-grown grafted apple during the winter. The clustering of differentially expressed genes (DEGs) and gene ontology enrichment indicated distinct expression dynamics in the two graft combinations, which supports the dependency of scion cold tolerance on the rootstock genotypes. We identified 544 potentially mobile mRNAs of DEGs showing highly-correlated seasonal dynamics between scion and rootstock. The mobility of a subset of 544 mRNAs was validated by translocated genome-wide variants and the measurements of selected RNA mobility in tobacco and Arabidopsis. We detected orthologous genes of potentially mobile mRNAs in Arabidopsis thaliana, which belong to cold regulatory networks with RNA mobility. Together, our study provides a comprehensive insight into gene interactions and signal exchange between scion and rootstock responsive to cold. This will serve for future research to enhance cold tolerance of grafted tree crops.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
