

Targeting AAV vectors to the central nervous system by engineering capsid-receptor interactions that enable crossing of the blood-brain barrier
- PMID: 37467291
- PMCID: PMC10355383
- DOI: 10.1371/journal.pbio.3002112
Targeting AAV vectors to the central nervous system by engineering capsid-receptor interactions that enable crossing of the blood-brain barrier
Abstract
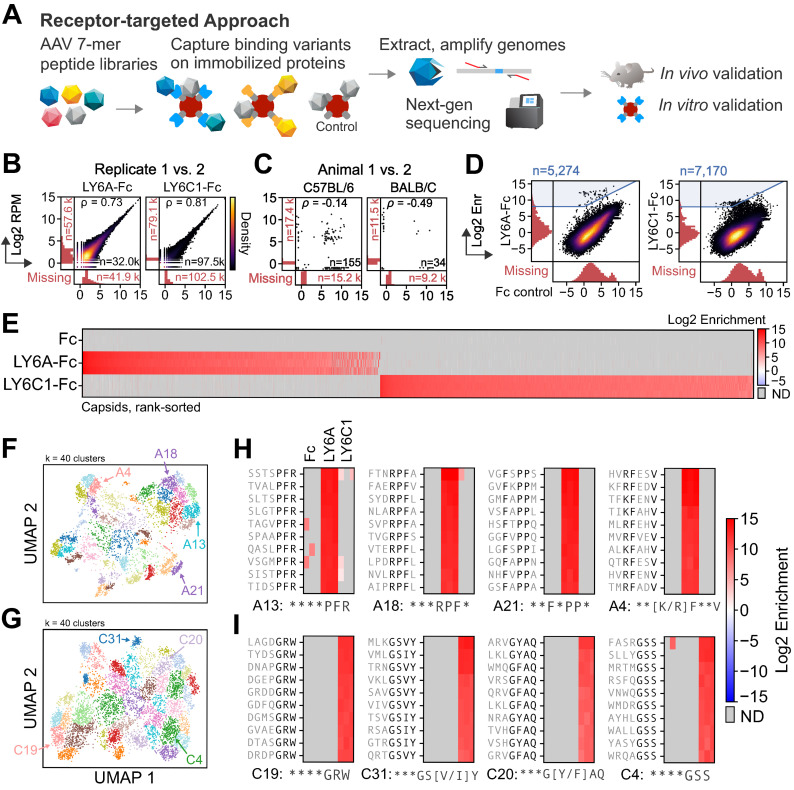
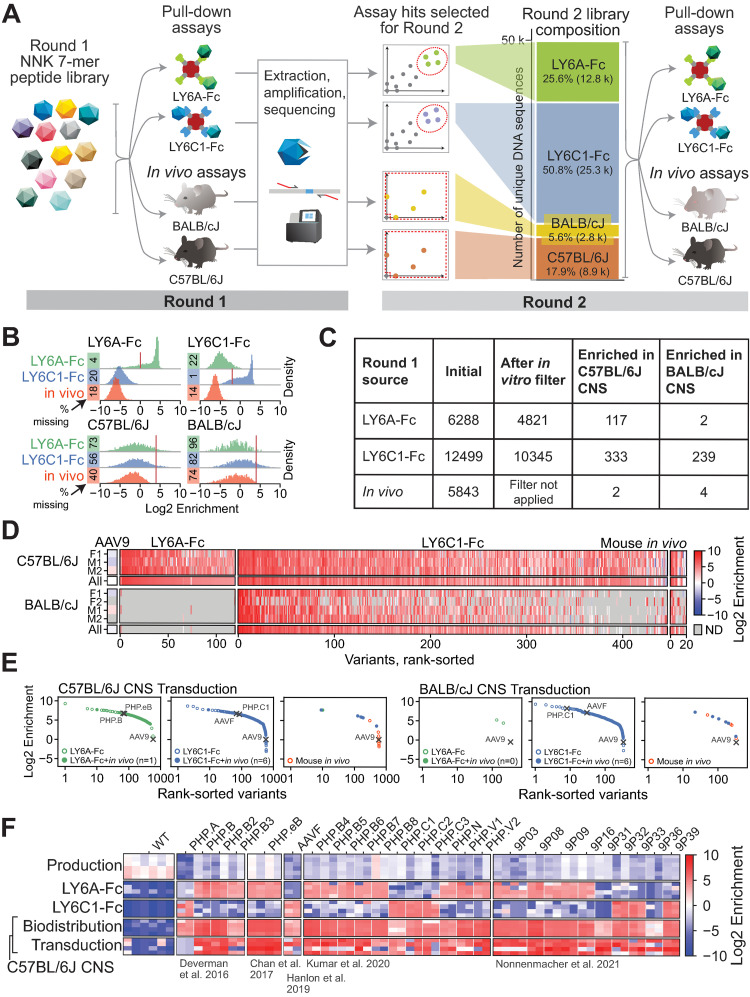
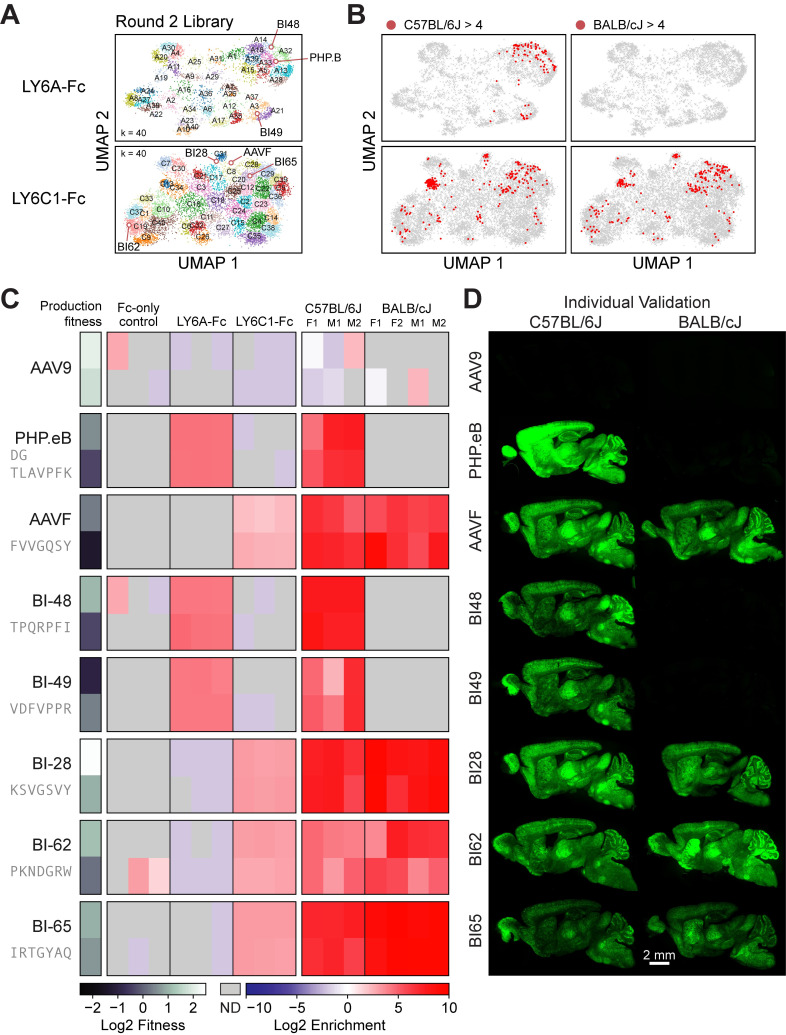
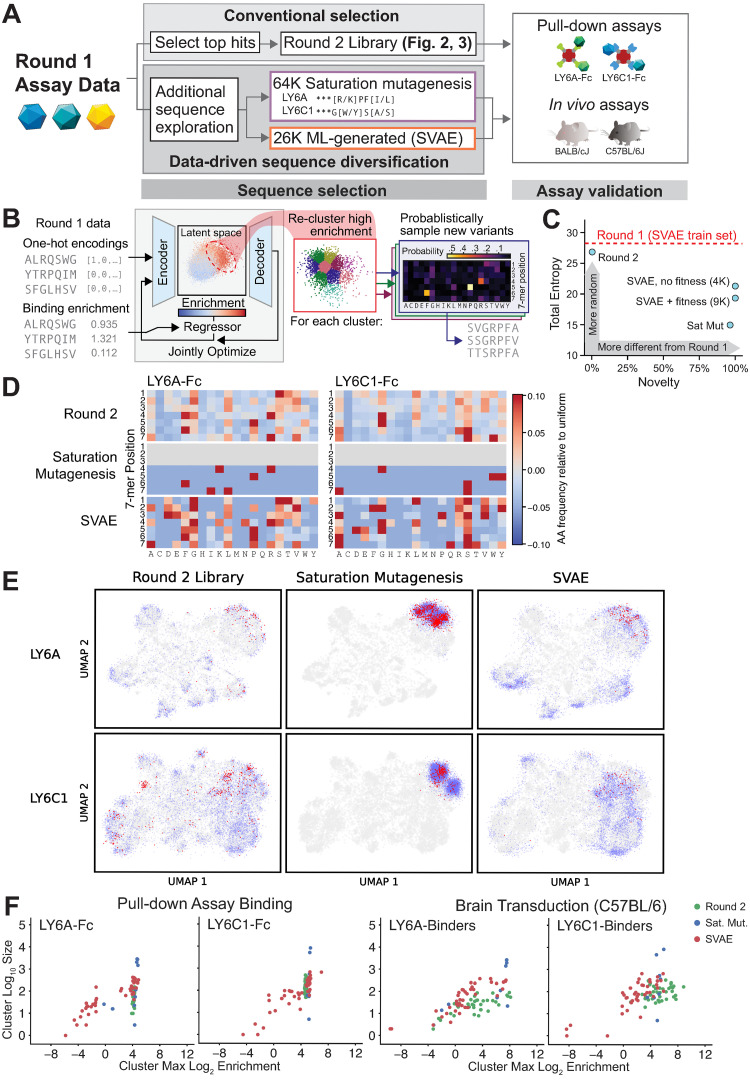
Viruses have evolved the ability to bind and enter cells through interactions with a wide variety of cell macromolecules. We engineered peptide-modified adeno-associated virus (AAV) capsids that transduce the brain through the introduction of de novo interactions with 2 proteins expressed on the mouse blood-brain barrier (BBB), LY6A or LY6C1. The in vivo tropisms of these capsids are predictable as they are dependent on the cell- and strain-specific expression of their target protein. This approach generated hundreds of capsids with dramatically enhanced central nervous system (CNS) tropisms within a single round of screening in vitro and secondary validation in vivo thereby reducing the use of animals in comparison to conventional multi-round in vivo selections. The reproducible and quantitative data derived via this method enabled both saturation mutagenesis and machine learning (ML)-guided exploration of the capsid sequence space. Notably, during our validation process, we determined that nearly all published AAV capsids that were selected for their ability to cross the BBB in mice leverage either the LY6A or LY6C1 protein, which are not present in primates. This work demonstrates that AAV capsids can be directly targeted to specific proteins to generate potent gene delivery vectors with known mechanisms of action and predictable tropisms.
Copyright: © 2023 Huang et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: BED is a scientific founder at Apertura Gene Therapy and a scientific advisory board member at Tevard Biosciences. BED, QH, KYC, and FEE are named inventors on patent applications filed by the Broad Institute of MIT and Harvard related to this study. Remaining authors declare that they have no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
