

Domains and residues of the Saccharomyces cerevisiae hnRNP protein Hrp1 important for transcriptional autoregulation and noncoding RNA termination
- PMID: 37467478
- PMCID: PMC10471224
- DOI: 10.1093/genetics/iyad134
Domains and residues of the Saccharomyces cerevisiae hnRNP protein Hrp1 important for transcriptional autoregulation and noncoding RNA termination
Abstract
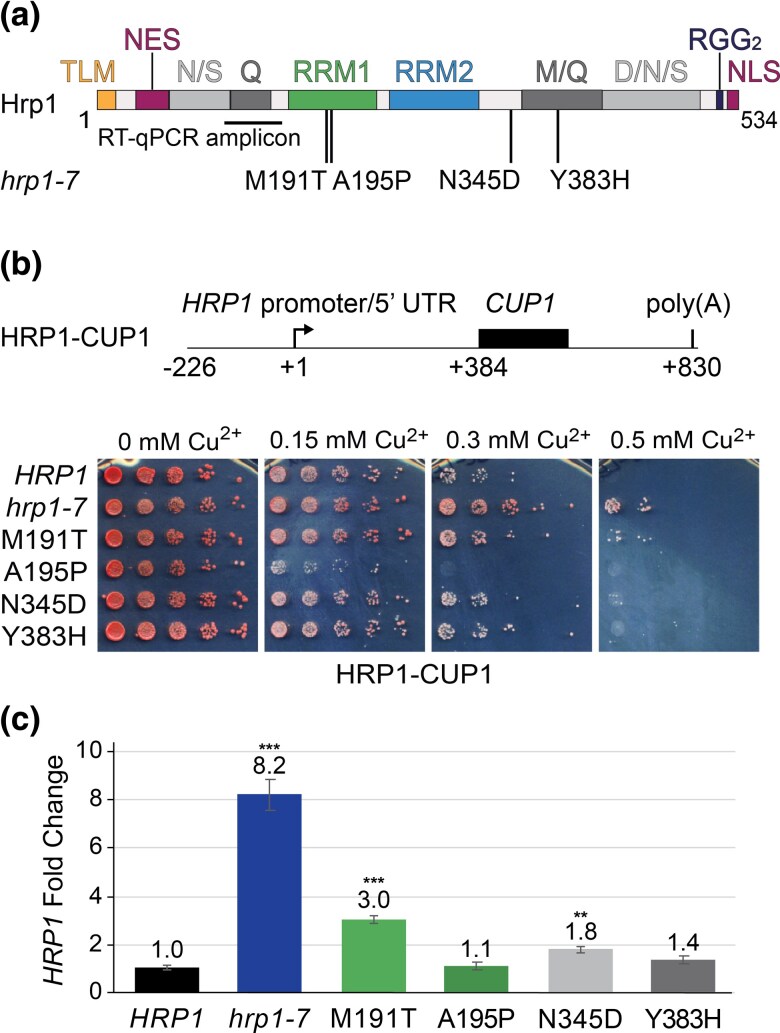
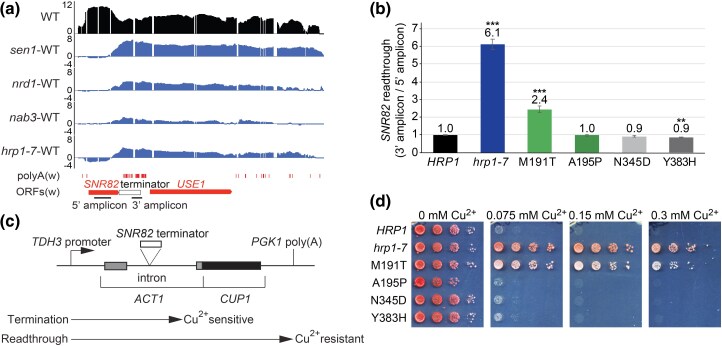
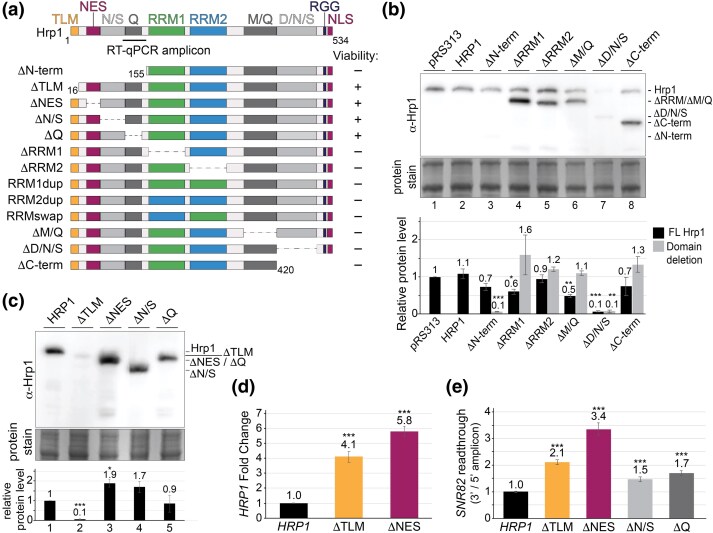
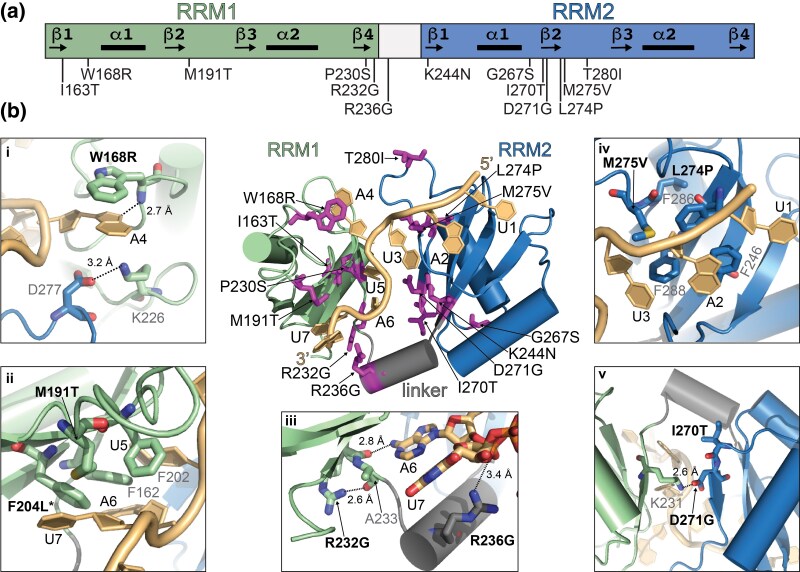
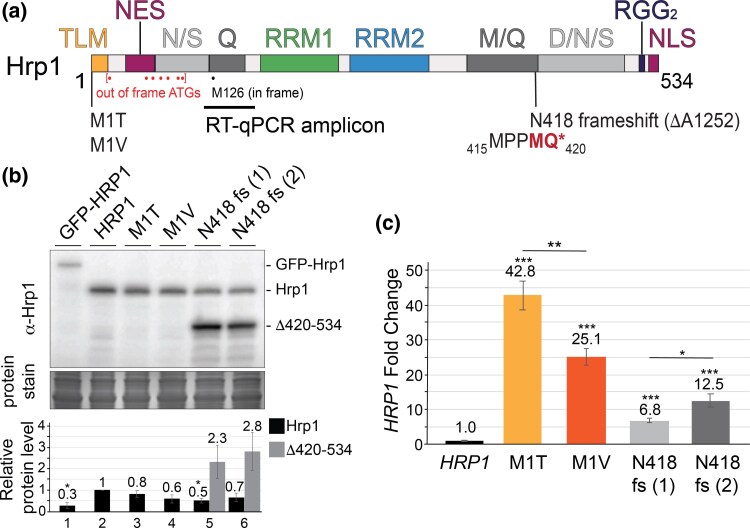
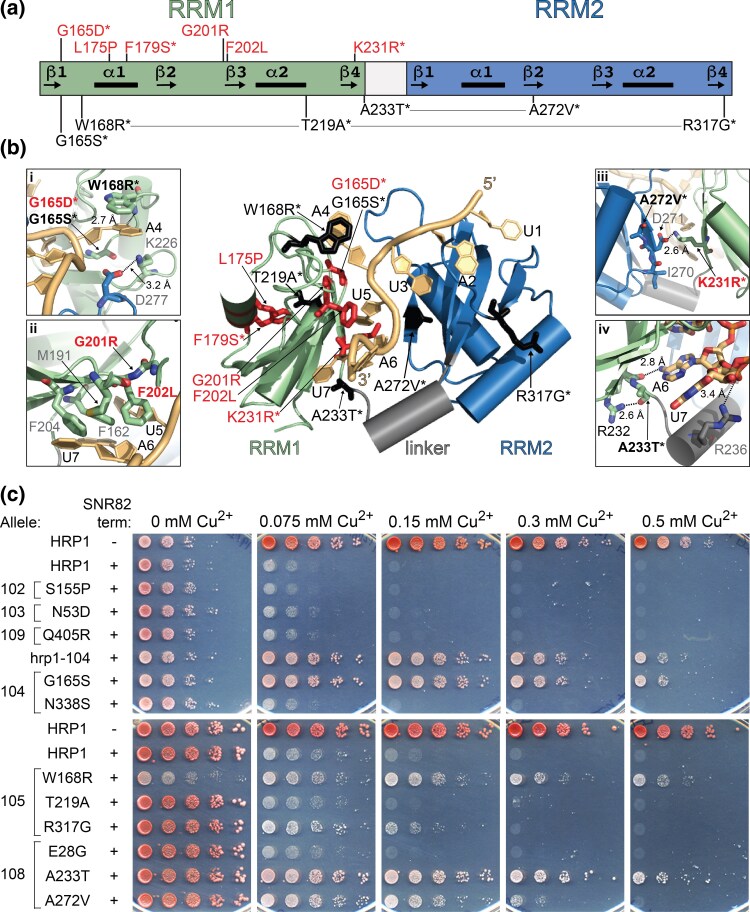
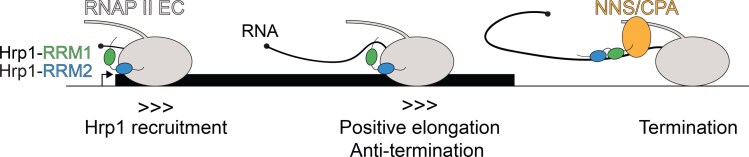
Proteins that bind the nascent transcript exiting RNA polymerase II can regulate transcription elongation. The essential Saccharomyces cerevisiae hnRNP protein Hrp1 is one such protein and participates in both cleavage and polyadenylation-coupled and Nrd1-Nab3-Sen1-dependent RNA polymerase II termination. Prior evidence that Hrp1 is a positive RNA polymerase II elongation factor suggests that its release from the elongation complex promotes termination. Here we report the effects of deletions and substitutions in Hrp1 on its autoregulation via an Nrd1-Nab3-Sen1-dependent transcription attenuator in the 5'-UTR of its mRNA and on the function of an Hrp1-dependent Nrd1-Nab3-Sen1 terminator in the SNR82 snoRNA gene. Deletion of either of two central RNA recognition motifs or either of the flanking low-sequence complexity domains is lethal. Smaller, viable deletions in the amino-terminal low-sequence complexity domain cause readthrough of both the HRP1 attenuator and SNR82 terminator. Substitutions that cause readthrough localized mostly to the RNA recognition motifs, although not always to the RNA-binding face. We found that autoregulation of Hrp1 mRNA synthesis is surprisingly robust, overcoming the expected lethal effects of the start codon and frameshift mutations via overexpression of the mRNA up to 40-fold. Our results suggest a model in which binding of attenuator or terminator elements in the nascent transcript by RNA recognition motifs 1 and 2 disrupts interactions between RNA recognition motif 2 and the RNA polymerase II elongation complex, increasing its susceptibility to termination.
Keywords: NNS termination; RNA polymerase II; hnRNP proteins; transcription; transcription termination; yeast.
© The Author(s) 2023. Published by Oxford University Press on behalf of The Genetics Society of America. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Conflict of interest statement
Conflicts of interest: The author(s) declare no conflict of interest.
Figures
Similar articles
-
RNA polymerase II transcription attenuation at the yeast DNA repair gene DEF1 is biologically significant and dependent on the Hrp1 RNA-recognition motif.G3 (Bethesda). 2023 Jan 12;13(1):jkac292. doi: 10.1093/g3journal/jkac292. G3 (Bethesda). 2023. PMID: 36315099 Free PMC article.
-
RNA Polymerase II Transcription Attenuation at the Yeast DNA Repair Gene, DEF1, Involves Sen1-Dependent and Polyadenylation Site-Dependent Termination.G3 (Bethesda). 2018 May 31;8(6):2043-2058. doi: 10.1534/g3.118.200072. G3 (Bethesda). 2018. PMID: 29686108 Free PMC article.
-
A network of interdependent molecular interactions describes a higher order Nrd1-Nab3 complex involved in yeast transcription termination.J Biol Chem. 2013 Nov 22;288(47):34158-34167. doi: 10.1074/jbc.M113.516765. Epub 2013 Oct 7. J Biol Chem. 2013. PMID: 24100036 Free PMC article.
-
The Nrd1-Nab3-Sen1 transcription termination complex from a structural perspective.Biochem Soc Trans. 2023 Jun 28;51(3):1257-1269. doi: 10.1042/BST20221418. Biochem Soc Trans. 2023. PMID: 37222282 Free PMC article. Review.
-
Termination of Transcription of Short Noncoding RNAs by RNA Polymerase II.Annu Rev Biochem. 2015;84:381-404. doi: 10.1146/annurev-biochem-060614-034457. Epub 2015 Mar 26. Annu Rev Biochem. 2015. PMID: 25747400 Free PMC article. Review.
Cited by
-
Mutations in yeast Pcf11, a conserved protein essential for mRNA 3' end processing and transcription termination, elicit the Environmental Stress Response.Genetics. 2024 Feb 7;226(2):iyad199. doi: 10.1093/genetics/iyad199. Genetics. 2024. PMID: 37967370 Free PMC article.
-
Substitutions in RNA-binding protein Hrp1 map a potential interaction surface with the yeast RNA polymerase II elongation complex.bioRxiv [Preprint]. 2025 May 7:2025.05.07.652672. doi: 10.1101/2025.05.07.652672. bioRxiv. 2025. PMID: 40654929 Free PMC article. Preprint.
References
-
- Arai T, Hasegawa M, Akiyama H, Ikeda K, Nonaka T, Mori H, Mann D, Tsuchiya K, Yoshida M, Hashizume Y, et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem Biophys Res Commun. 2006;351(3):602–611. doi: 10.1016/j.bbrc.2006.10.093. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
