

Ethylene-mediated metabolic priming increases photosynthesis and metabolism to enhance plant growth and stress tolerance
- PMID: 37469928
- PMCID: PMC10353721
- DOI: 10.1093/pnasnexus/pgad216
Ethylene-mediated metabolic priming increases photosynthesis and metabolism to enhance plant growth and stress tolerance
Abstract
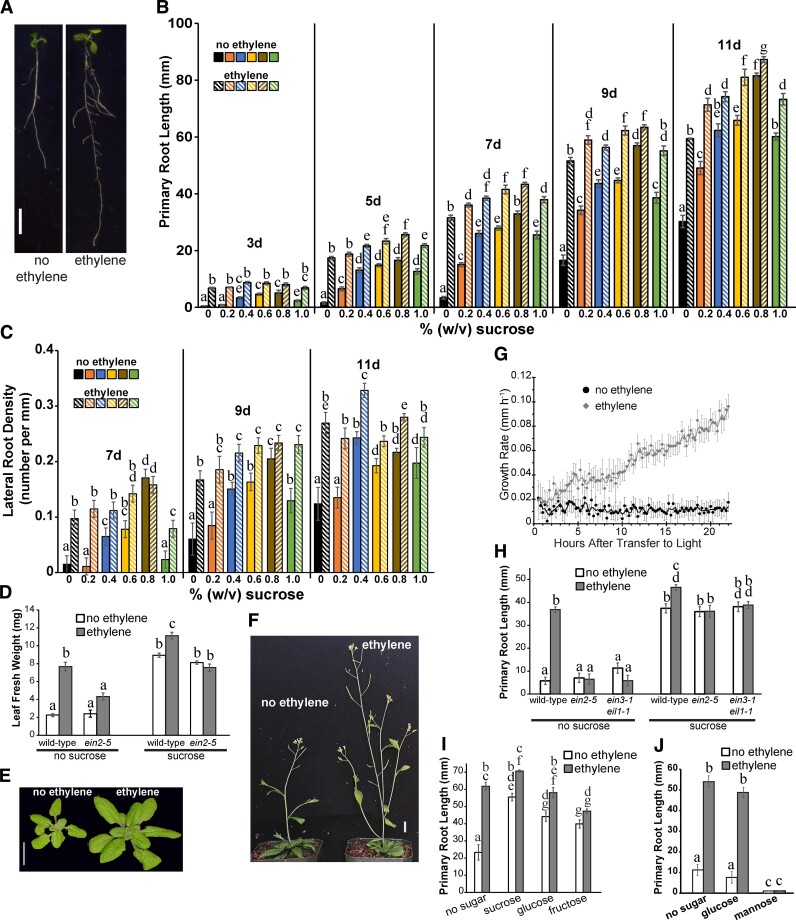
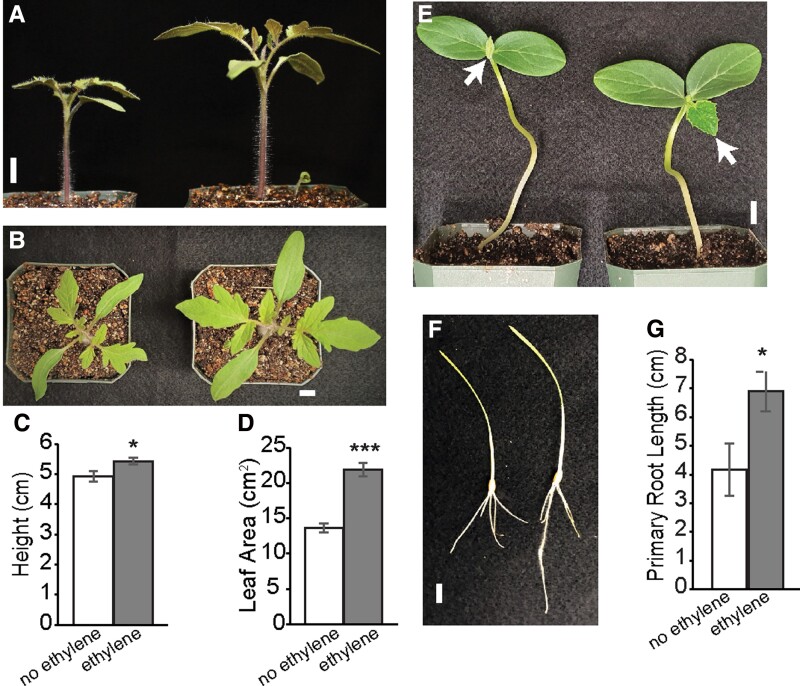
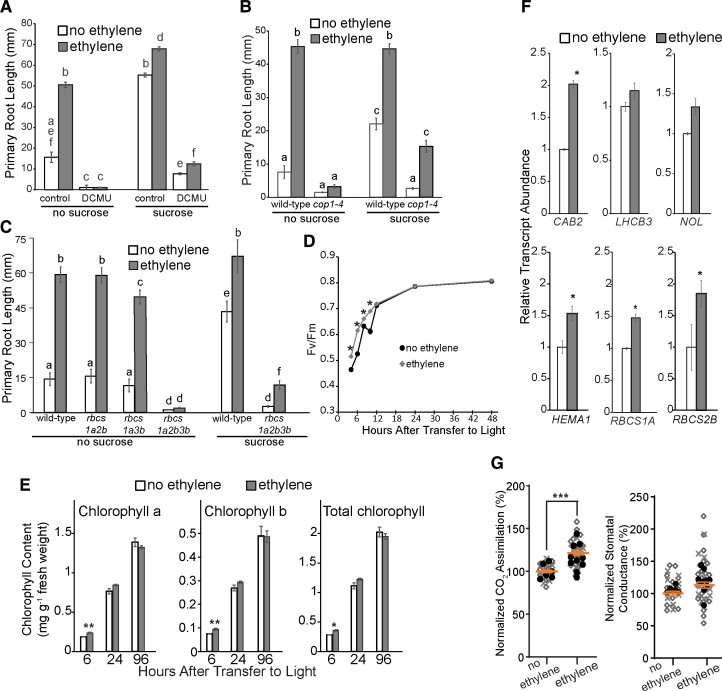
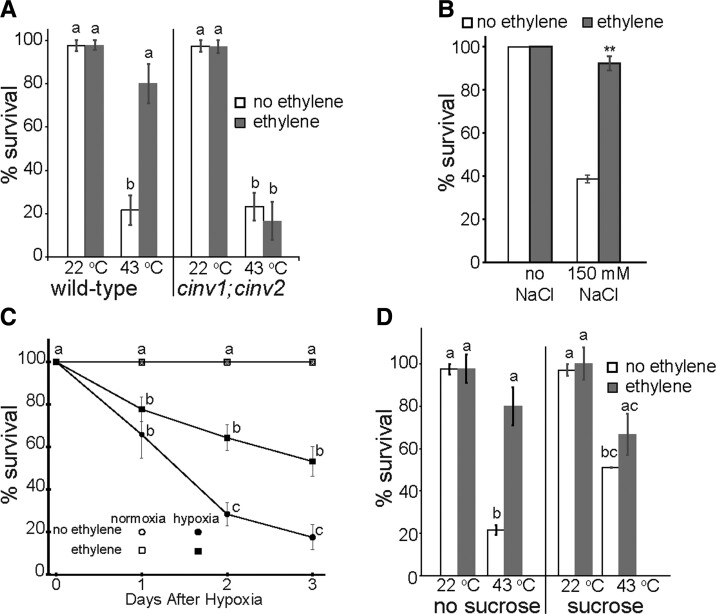
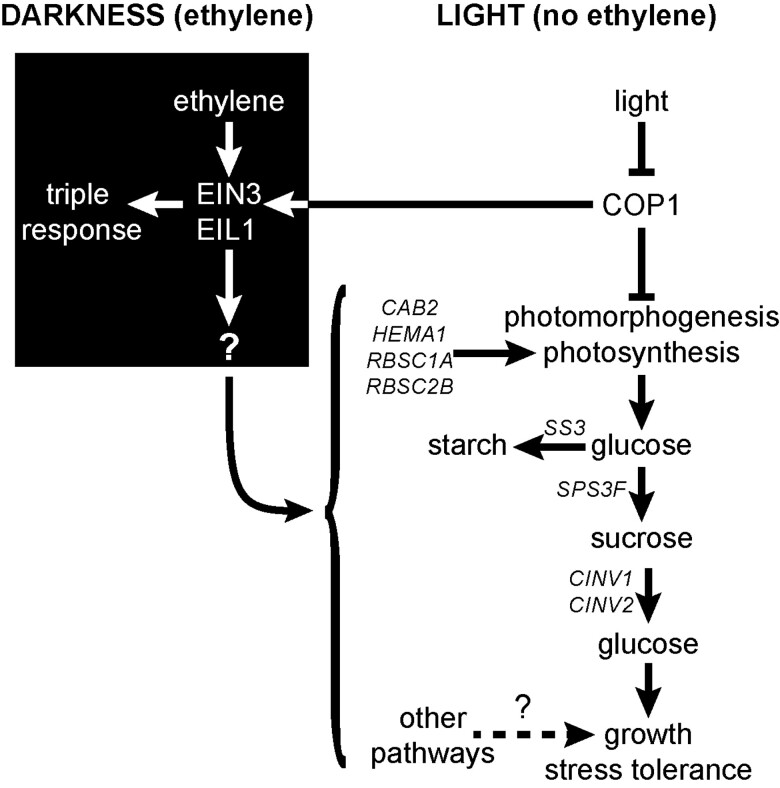
Enhancing crop yields is a major challenge because of an increasing human population, climate change, and reduction in arable land. Here, we demonstrate that long-lasting growth enhancement and increased stress tolerance occur by pretreatment of dark grown Arabidopsis seedlings with ethylene before transitioning into light. Plants treated this way had longer primary roots, more and longer lateral roots, and larger aerial tissue and were more tolerant to high temperature, salt, and recovery from hypoxia stress. We attributed the increase in plant growth and stress tolerance to ethylene-induced photosynthetic-derived sugars because ethylene pretreatment caused a 23% increase in carbon assimilation and increased the levels of glucose (266%), sucrose/trehalose (446%), and starch (87%). Metabolomic and transcriptomic analyses several days posttreatment showed a significant increase in metabolic processes and gene transcripts implicated in cell division, photosynthesis, and carbohydrate metabolism. Because of this large effect on metabolism, we term this "ethylene-mediated metabolic priming." Reducing photosynthesis with inhibitors or mutants prevented the growth enhancement, but this was partially rescued by exogenous sucrose, implicating sugars in this growth phenomenon. Additionally, ethylene pretreatment increased the levels of CINV1 and CINV2 encoding invertases that hydrolyze sucrose, and cinv1;cinv2 mutants did not respond to ethylene pretreatment with increased growth indicating increased sucrose breakdown is critical for this trait. A model is proposed where ethylene-mediated metabolic priming causes long-term increases in photosynthesis and carbohydrate utilization to increase growth. These responses may be part of the natural development of seedlings as they navigate through the soil to emerge into light.
Keywords: ethylene; growth; metabolism; photosynthesis; stress tolerance.
© The Author(s) 2023. Published by Oxford University Press on behalf of National Academy of Sciences.
Figures

References
-
- Abeles F, Morgan P, Saltveit MJ. 1992. Ethylene in plant biology. 2nd ed. San Diego, CA: Academic Press. p. 414.
-
- Khan NA, Ferrante A, Munné-Bosch S. 2022. The plant hormone ethylene: stress acclimation and agricultural applications. Cambridge (MA): Elsevier. p. 248.
-
- Bleecker AB, Estelle MA, Somerville C, Kende H. 1988. Insensitivity to ethylene conferred by a dominant mutation in Arabidopsis thaliana. Science 241:1086–1089. - PubMed
-
- Chang C, Kwok SF, Bleecker AB, Meyerowitz EM. 1993. Arabidopsis ethylene-response gene ETR1: similarity of product to two-component regulators. Science 262:539–544. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
