

A dominant-negative avirulence effector of the barley powdery mildew fungus provides mechanistic insight into barley MLA immune receptor activation
- PMID: 37474129
- PMCID: PMC10540733
- DOI: 10.1093/jxb/erad285
A dominant-negative avirulence effector of the barley powdery mildew fungus provides mechanistic insight into barley MLA immune receptor activation
Abstract
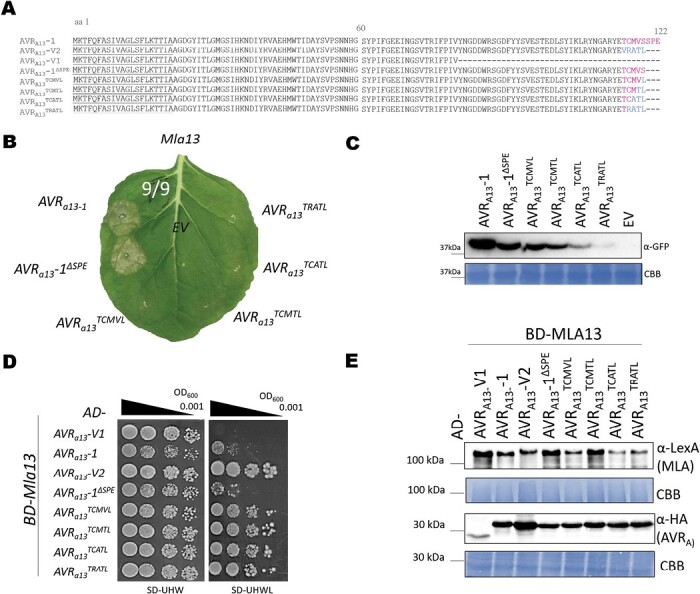
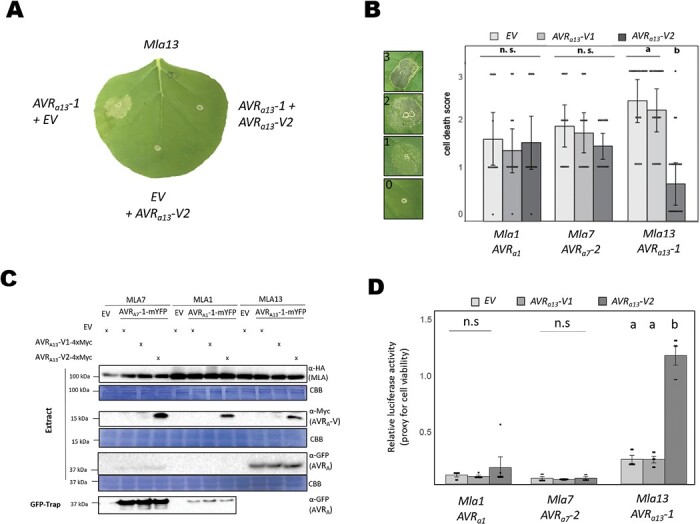
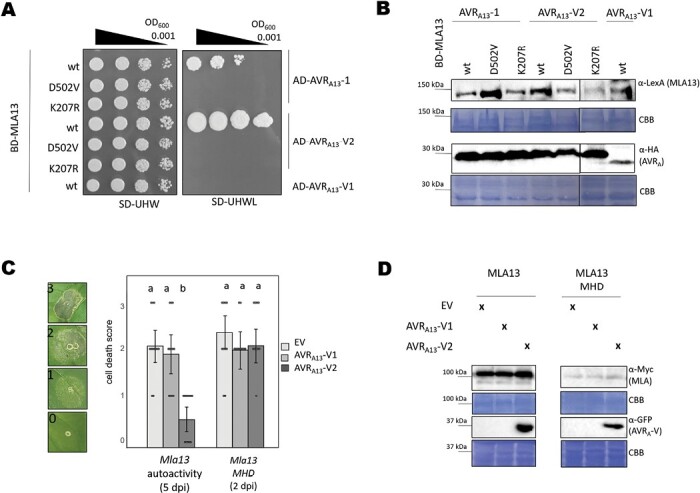
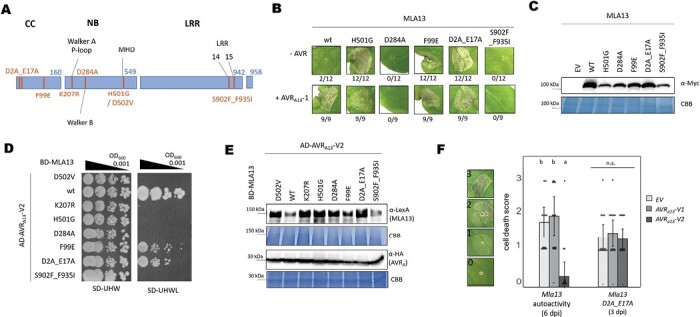
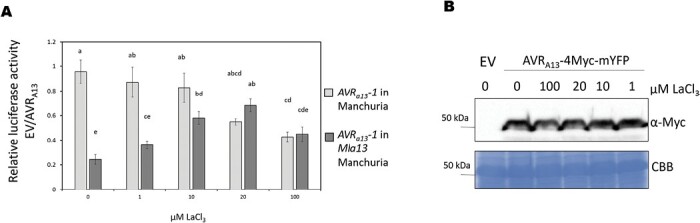
Nucleotide-binding leucine-rich repeat receptors (NLRs) recognize pathogen effectors to mediate plant disease resistance often involving host cell death. Effectors escape NLR recognition through polymorphisms, allowing the pathogen to proliferate on previously resistant host plants. The powdery mildew effector AVRA13-1 is recognized by the barley NLR MLA13 and activates host cell death. We demonstrate here that a virulent form of AVRA13, called AVRA13-V2, escapes MLA13 recognition by substituting a serine for a leucine residue at the C-terminus. Counterintuitively, this substitution in AVRA13-V2 resulted in an enhanced MLA13 association and prevented the detection of AVRA13-1 by MLA13. Therefore, AVRA13-V2 is a dominant-negative form of AVRA13 and has probably contributed to the breakdown of Mla13 resistance. Despite this dominant-negative activity, AVRA13-V2 failed to suppress host cell death mediated by the MLA13 autoactive MHD variant. Neither AVRA13-1 nor AVRA13-V2 interacted with the MLA13 autoactive variant, implying that the binding moiety in MLA13 that mediates association with AVRA13-1 is altered after receptor activation. We also show that mutations in the MLA13 coiled-coil domain, which were thought to impair Ca2+ channel activity and NLR function, instead resulted in MLA13 autoactive cell death. Our results constitute an important step to define intermediate receptor conformations during NLR activation.
Keywords: Blumeria graminis; AVR; MLA; Mildew Locus A; NLR; barley; cell death; fungal effector; powdery mildew; resistance.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Conflict of interest statement
No conflict of interest declared.
Figures
References
-
- Bauer S, Yu D, Lawson AW, Saur IM, Frantzeskakis L, Kracher B, Logemann E, Chai J, Maekawa T, Schulze-Lefert P.. 2021. The leucine-rich repeats in allelic barley MLA immune receptors define specificity towards sequence-unrelated powdery mildew avirulence effectors with a predicted common RNase-like fold. PLoS Pathogens 17, e1009223. - PMC - PubMed
-
- Bendahmane A, Farnham G, Moffett P, Baulcombe DC.. 2002. Constitutive gain-of-function mutants in a nucleotide binding site-leucine rich repeat protein encoded at the Rx locus of potato. The Plant Journal 32, 195–204. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
