

A maize epimerase modulates cell wall synthesis and glycosylation during stomatal morphogenesis
- PMID: 37474494
- PMCID: PMC10359280
- DOI: 10.1038/s41467-023-40013-6
A maize epimerase modulates cell wall synthesis and glycosylation during stomatal morphogenesis
Abstract
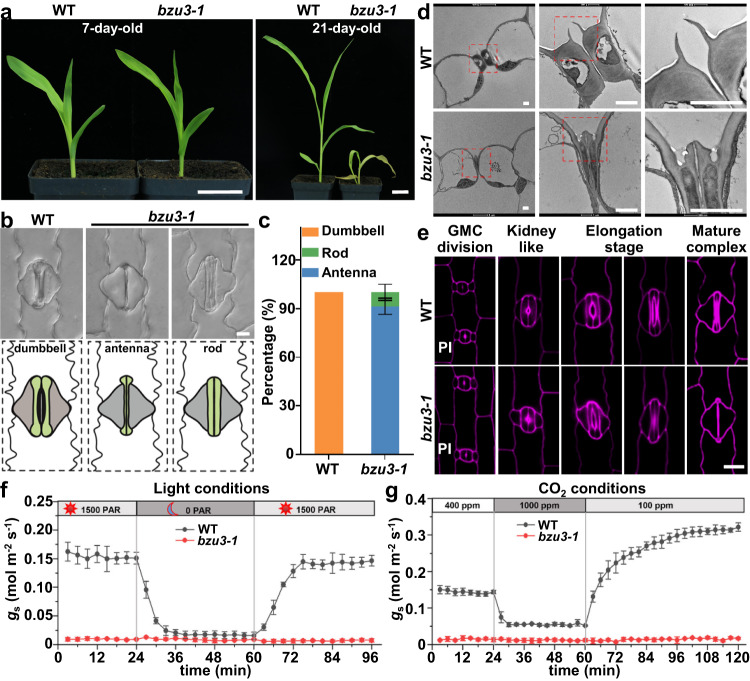
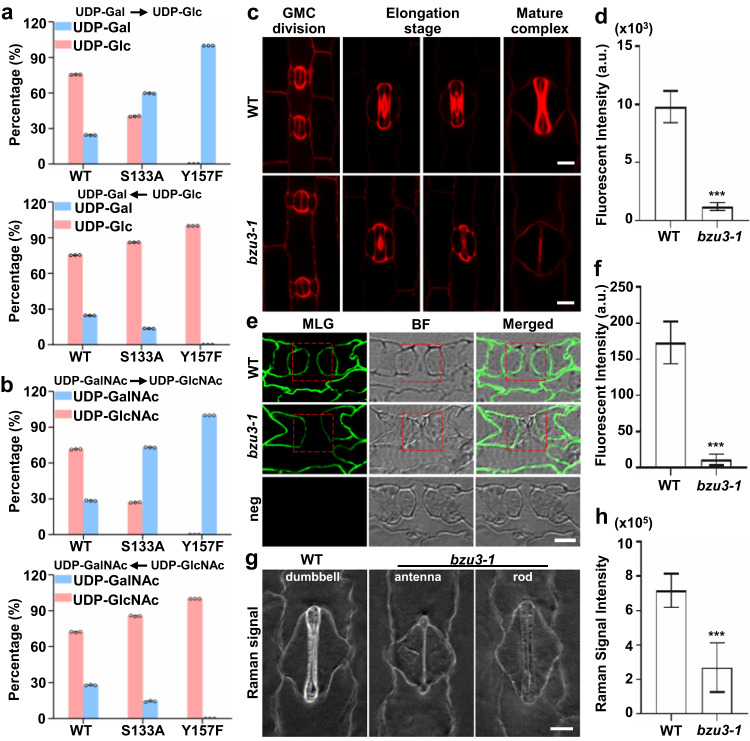
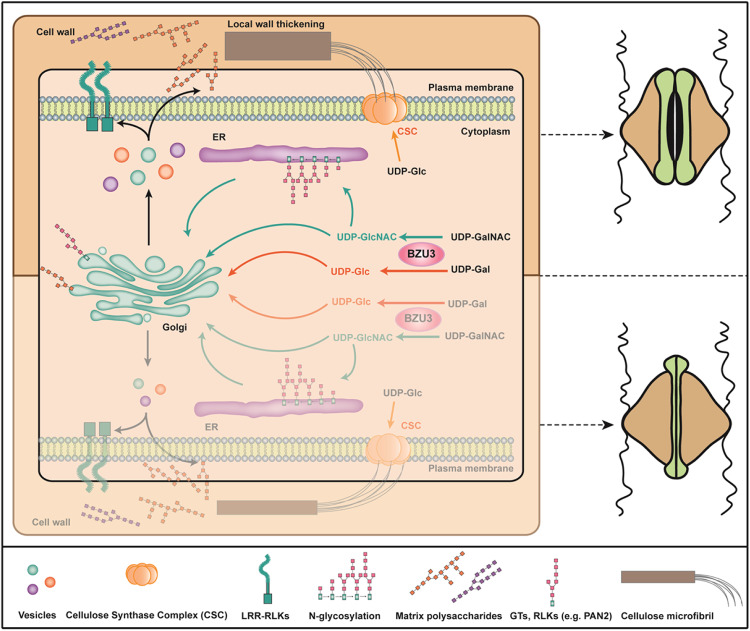
The unique dumbbell-shape of grass guard cells (GCs) is controlled by their cell walls which enable their rapid responses to the environment. The molecular mechanisms regulating the synthesis and assembly of GC walls are as yet unknown. Here we have identified BZU3, a maize gene encoding UDP-glucose 4-epimerase that regulates the supply of UDP-glucose during GC wall synthesis. The BZU3 mutation leads to significant decreases in cellular UDP-glucose levels. Immunofluorescence intensities reporting levels of cellulose and mixed-linkage glucans are reduced in the GCs, resulting in impaired local wall thickening. BZU3 also catalyzes the epimerization of UDP-N-acetylgalactosamine to UDP-N-acetylglucosamine, and the BZU3 mutation affects N-glycosylation of proteins that may be involved in cell wall synthesis and signaling. Our results suggest that the spatiotemporal modulation of BZU3 plays a dual role in controlling cell wall synthesis and glycosylation via controlling UDP-glucose/N-acetylglucosamine homeostasis during stomatal morphogenesis. These findings provide insights into the mechanisms controlling formation of the unique morphology of grass stomata.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Ezquer I, Salameh I, Colombo L, Kalaitzis P. Plant cell walls tackling climate change: insights into plant cell wall remodeling, its regulation, and biotechnological strategies to improve crop adaptations and photosynthesis in response to global warming. Plants (Basel) 2020;9:212. doi: 10.3390/plants9020212. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
