

ThPOK is a critical multifaceted regulator of myeloid lineage development
- PMID: 37474652
- PMCID: PMC10792516
- DOI: 10.1038/s41590-023-01549-3
ThPOK is a critical multifaceted regulator of myeloid lineage development
Abstract
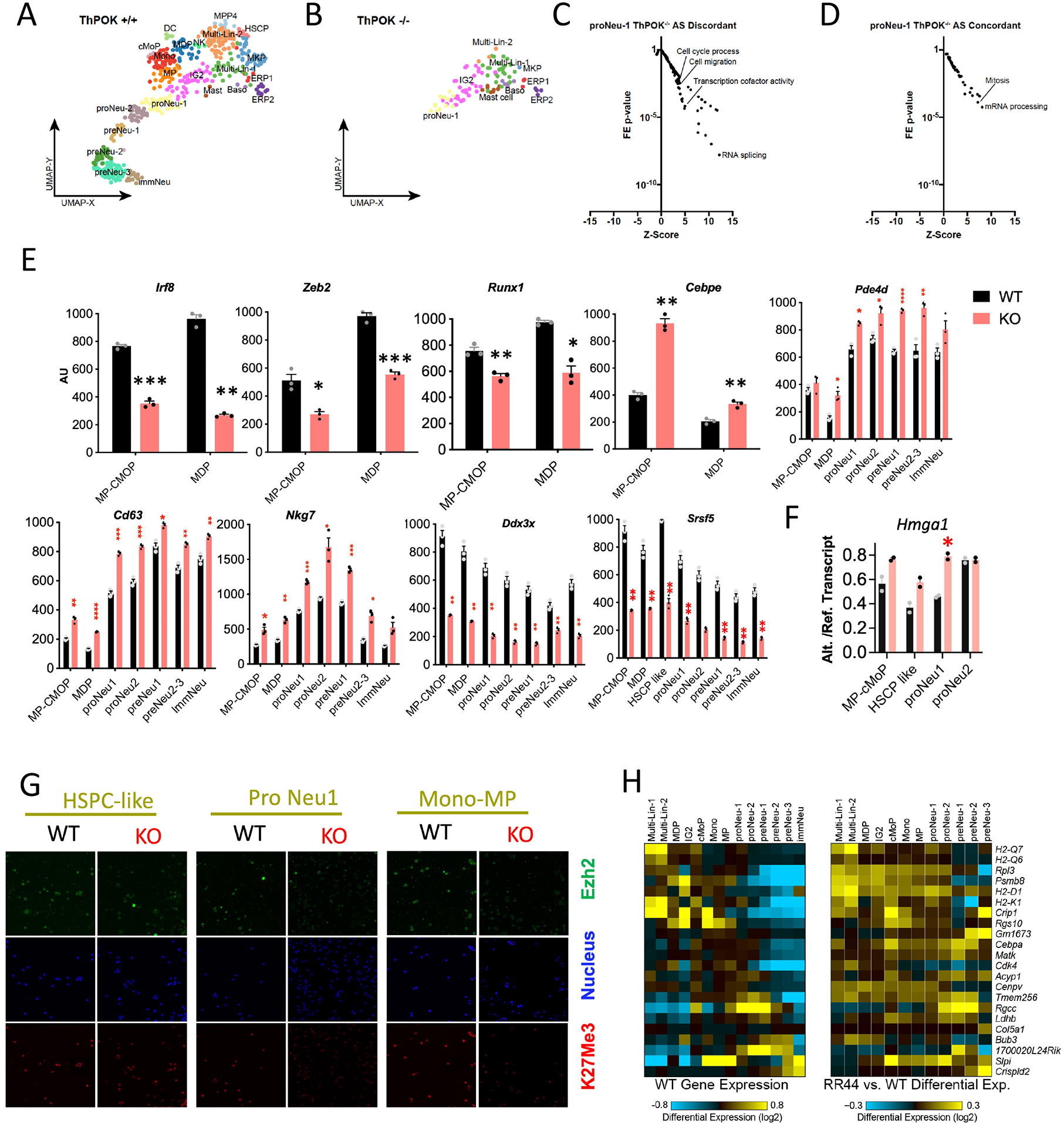
The transcription factor ThPOK (encoded by Zbtb7b) is well known for its role as a master regulator of CD4 lineage commitment in the thymus. Here, we report an unexpected and critical role of ThPOK as a multifaceted regulator of myeloid lineage commitment, differentiation and maturation. Using reporter and knockout mouse models combined with single-cell RNA-sequencing, progenitor transfer and colony assays, we show that ThPOK controls monocyte-dendritic cell versus granulocyte lineage production during homeostatic differentiation, and serves as a brake for neutrophil maturation in granulocyte lineage-specified cells through transcriptional regulation of lineage-specific transcription factors and RNA via altered messenger RNA splicing to reprogram intron retention.
© 2023. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Figures













References
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL122661/HL/NHLBI NIH HHS/United States
- R01 GM107179/GM/NIGMS NIH HHS/United States
- AI164333/AI/NIAID NIH HHS/United States
- R21 AI164333/AI/NIAID NIH HHS/United States
- GM107179/GM/NIGMS NIH HHS/United States
- AI068907/AI/NIAID NIH HHS/United States
- CA211479/CA/NCI NIH HHS/United States
- CA195356/CA/NCI NIH HHS/United States
- HL122661/HL/NHLBI NIH HHS/United States
- R01 CA226802/CA/NCI NIH HHS/United States
- P30 CA006927/CA/NCI NIH HHS/United States
- RC2 DK122376/DK/NIDDK NIH HHS/United States
- R01 AI068907/AI/NIAID NIH HHS/United States
- R21 CA195356/CA/NCI NIH HHS/United States
- R50 CA211479/CA/NCI NIH HHS/United States
- CA006927/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
