

Axonal energy metabolism, and the effects in aging and neurodegenerative diseases
- PMID: 37475056
- PMCID: PMC10357692
- DOI: 10.1186/s13024-023-00634-3
Axonal energy metabolism, and the effects in aging and neurodegenerative diseases
Abstract
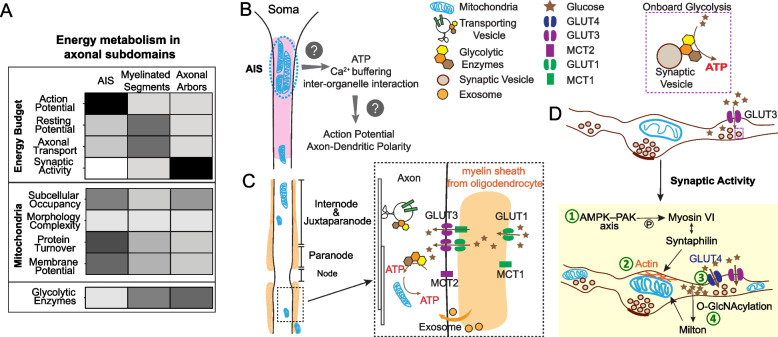
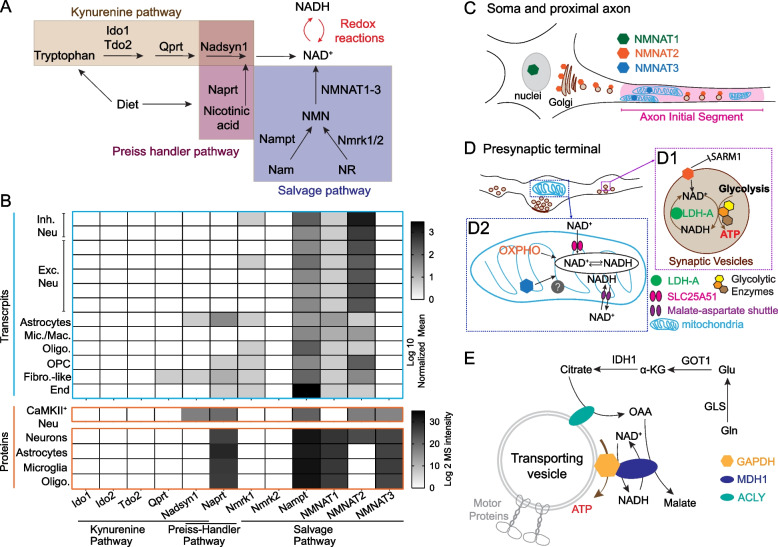
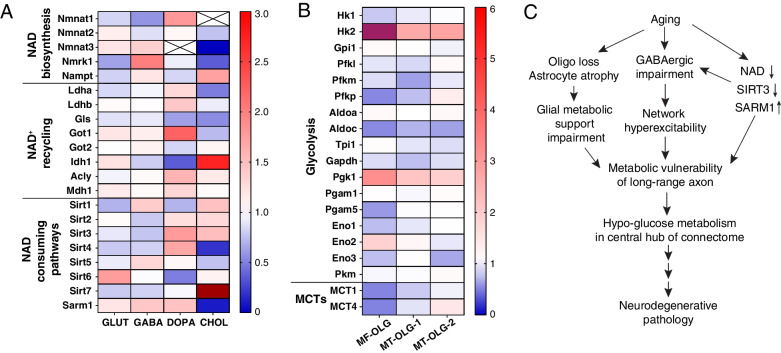
Human studies consistently identify bioenergetic maladaptations in brains upon aging and neurodegenerative disorders of aging (NDAs), such as Alzheimer's disease, Parkinson's disease, Huntington's disease, and Amyotrophic lateral sclerosis. Glucose is the major brain fuel and glucose hypometabolism has been observed in brain regions vulnerable to aging and NDAs. Many neurodegenerative susceptible regions are in the topological central hub of the brain connectome, linked by densely interconnected long-range axons. Axons, key components of the connectome, have high metabolic needs to support neurotransmission and other essential activities. Long-range axons are particularly vulnerable to injury, neurotoxin exposure, protein stress, lysosomal dysfunction, etc. Axonopathy is often an early sign of neurodegeneration. Recent studies ascribe axonal maintenance failures to local bioenergetic dysregulation. With this review, we aim to stimulate research in exploring metabolically oriented neuroprotection strategies to enhance or normalize bioenergetics in NDA models. Here we start by summarizing evidence from human patients and animal models to reveal the correlation between glucose hypometabolism and connectomic disintegration upon aging/NDAs. To encourage mechanistic investigations on how axonal bioenergetic dysregulation occurs during aging/NDAs, we first review the current literature on axonal bioenergetics in distinct axonal subdomains: axon initial segments, myelinated axonal segments, and axonal arbors harboring pre-synaptic boutons. In each subdomain, we focus on the organization, activity-dependent regulation of the bioenergetic system, and external glial support. Second, we review the mechanisms regulating axonal nicotinamide adenine dinucleotide (NAD+) homeostasis, an essential molecule for energy metabolism processes, including NAD+ biosynthetic, recycling, and consuming pathways. Third, we highlight the innate metabolic vulnerability of the brain connectome and discuss its perturbation during aging and NDAs. As axonal bioenergetic deficits are developing into NDAs, especially in asymptomatic phase, they are likely exaggerated further by impaired NAD+ homeostasis, the high energetic cost of neural network hyperactivity, and glial pathology. Future research in interrogating the causal relationship between metabolic vulnerability, axonopathy, amyloid/tau pathology, and cognitive decline will provide fundamental knowledge for developing therapeutic interventions.
Keywords: Aging; Axonal bioenergetics; Axonopathy; Energy metabolism; Glucose; Glycolysis; Mitochondria; NAD; Neurodegeneration; Neuroprotection.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Erbsloh F, Bernsmeier A, Hillesheim H. The glucose consumption of the brain & its dependence on the liver. Arch Psychiatr Nervenkr Z Gesamte Neurol Psychiatr. 1958;196:611–626. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
