

The coevolutionary dynamics of cryptic female choice
- PMID: 37475752
- PMCID: PMC10355280
- DOI: 10.1093/evlett/qrad025
The coevolutionary dynamics of cryptic female choice
Abstract
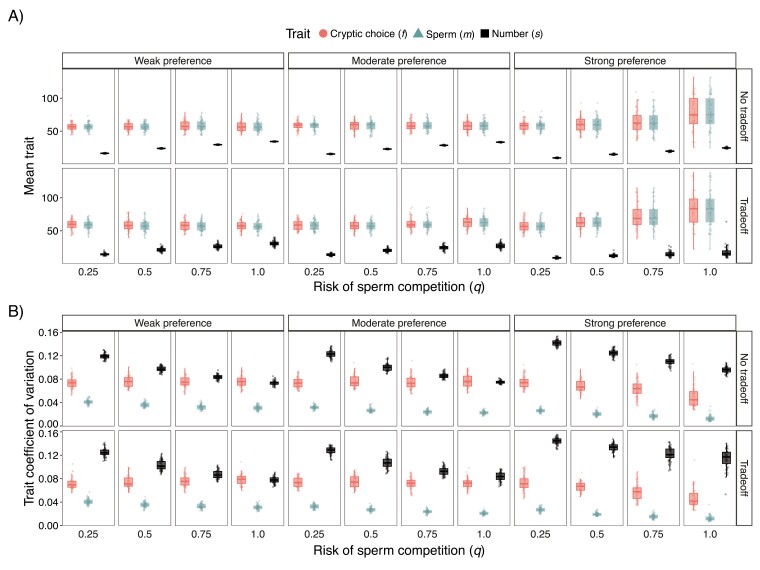
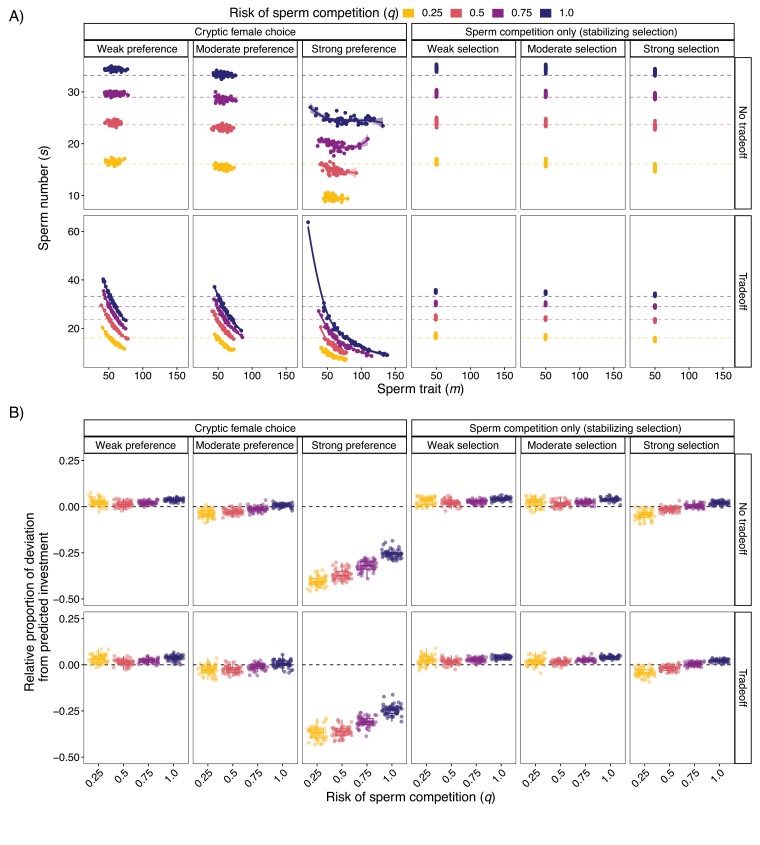
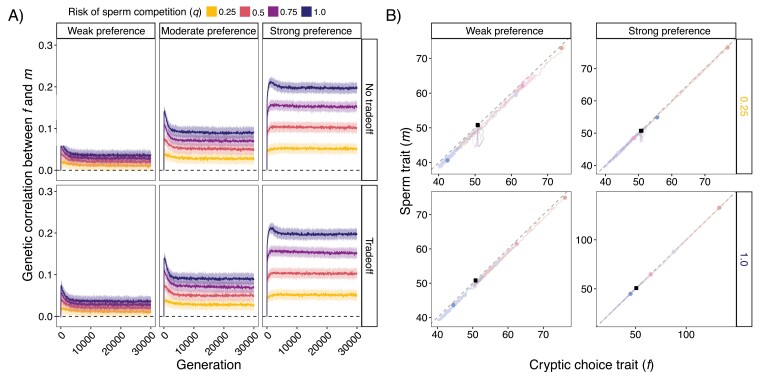
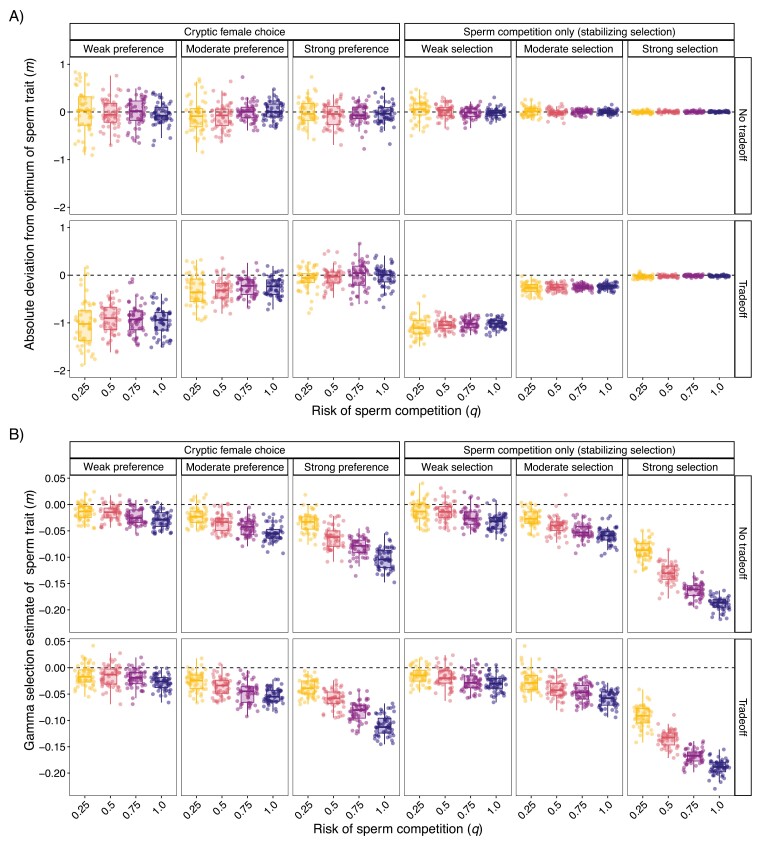
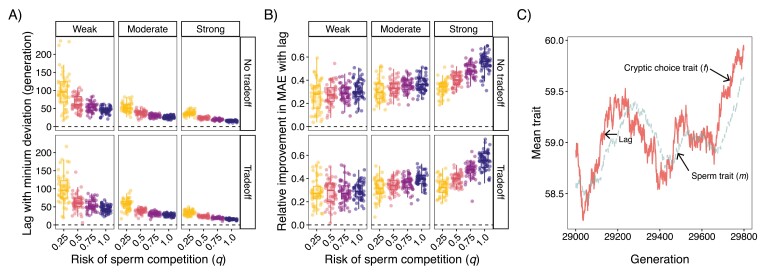
In contrast to sexual selection on traits that affect interactions between the sexes before mating, little theoretical research has focused on the coevolution of postmating traits via cryptic female choice (when females bias fertilization toward specific males). We used simulation models to ask (a) whether and, if so, how nondirectional cryptic female choice (female-by-male interactions in fertilization success) causes deviations from models that focus exclusively on male-mediated postmating processes, and (b) how the risk of sperm competition, the strength of cryptic female choice, and tradeoffs between sperm number and sperm traits interact to influence the coevolutionary dynamics between cryptic female choice and sperm traits. We found that incorporating cryptic female choice can result in males investing much less in their ejaculates than predicted by models with sperm competition only. We also found that cryptic female choice resulted in the evolution of genetic correlations between cryptic female choice and sperm traits, even when the strength of cryptic female choice was weak, and the risk of sperm competition was low. This suggests that cryptic female choice may be important even in systems with low multiple mating. These genetic correlations increased with the risk of sperm competition and as the strength of cryptic female choice increased. When the strength of cryptic female choice and risk of sperm competition was high, extreme codivergence of sperm traits and cryptic female choice preference occurred even when the sperm trait traded off with sperm number. We also found that male traits lagged behind the evolution of female traits; this lag decreased with increasing strength of cryptic female choice and risk of sperm competition. Overall, our results suggest that cryptic female choice deserves more attention theoretically and may be driving trait evolution in ways just beginning to be explored.
Keywords: Fisherian Runaway Selection; individual-based model; postcopulatory sexual selection; reproductive isolation; sperm competition.
© The Author(s) 2023. Published by Oxford University Press on behalf of The Society for the Study of Evolution (SSE) and European Society for Evolutionary Biology (ESEN).
Figures
References
-
- Bezanson, J., Edelman, A., Karpinski, S., & Shah, V. B. (2017). Julia: A fresh approach to numerical computing. SIAM Review, 59(1), 65–98. 10.1137/141000671 - DOI
