

Mechanical confinement promotes heat resistance of hepatocellular carcinoma via SP1/IL4I1/AHR axis
- PMID: 37478857
- PMCID: PMC10439175
- DOI: 10.1016/j.xcrm.2023.101128
Mechanical confinement promotes heat resistance of hepatocellular carcinoma via SP1/IL4I1/AHR axis
Abstract
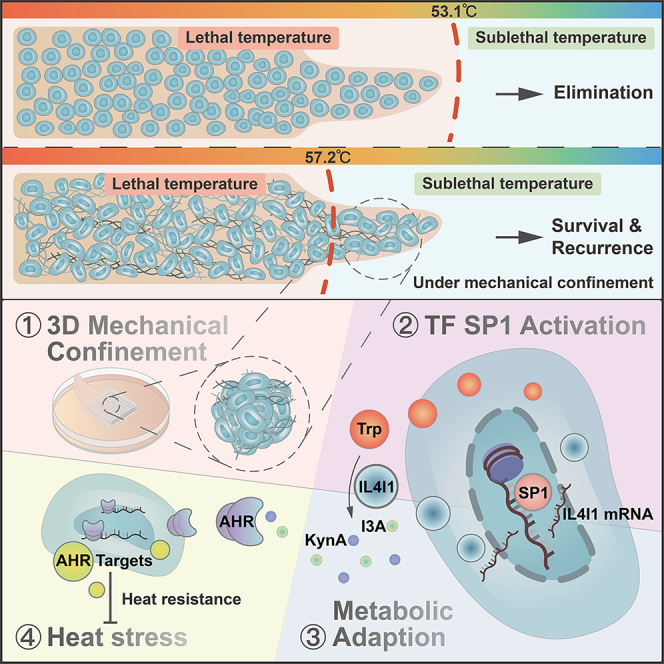
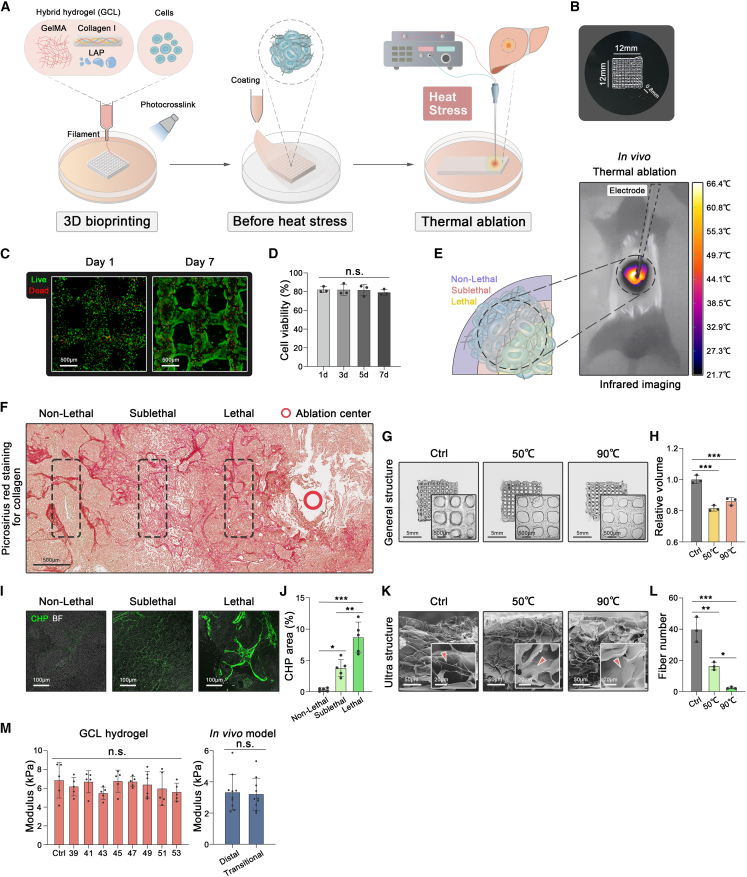
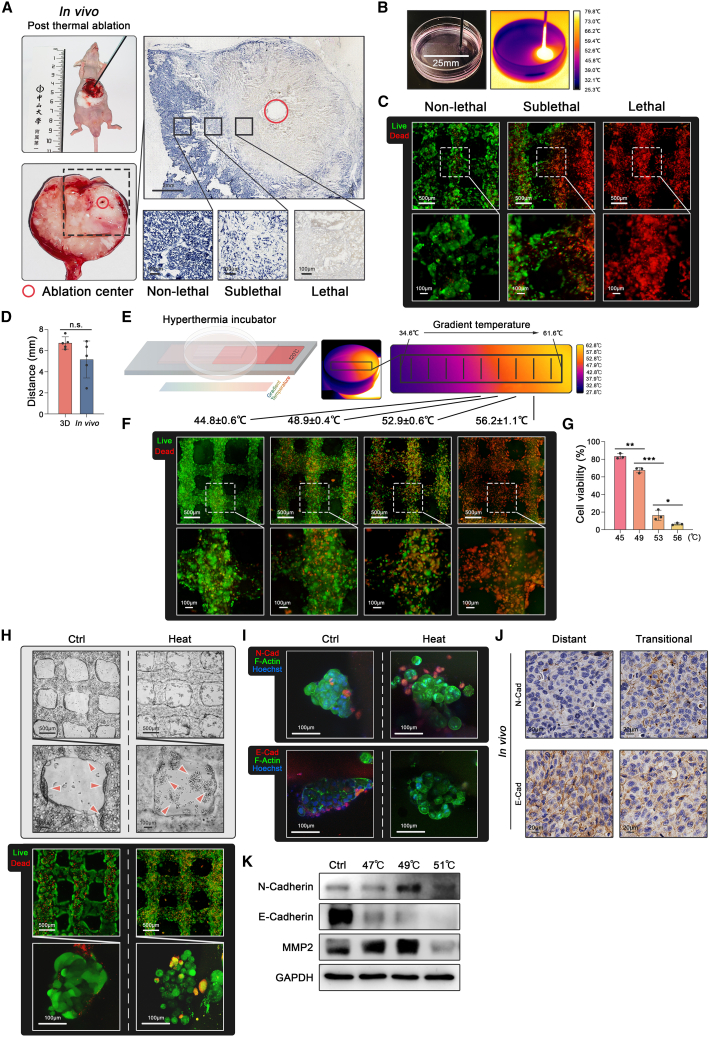
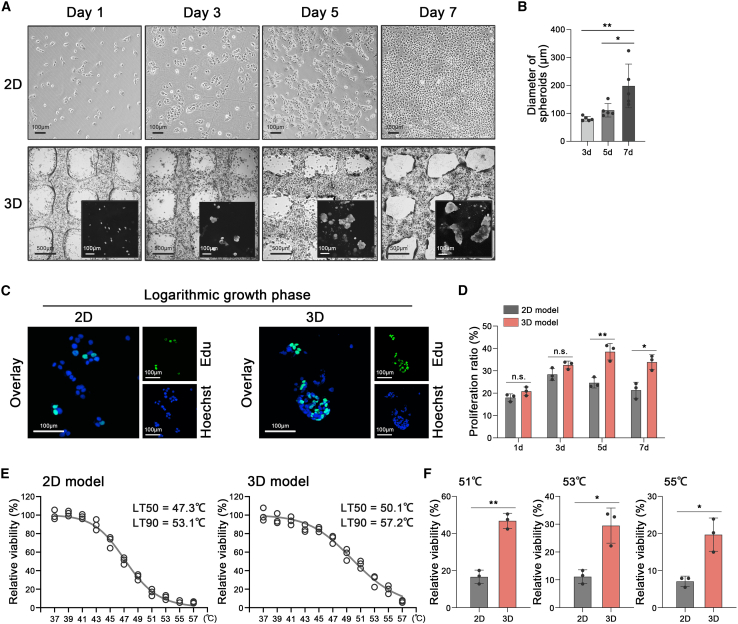
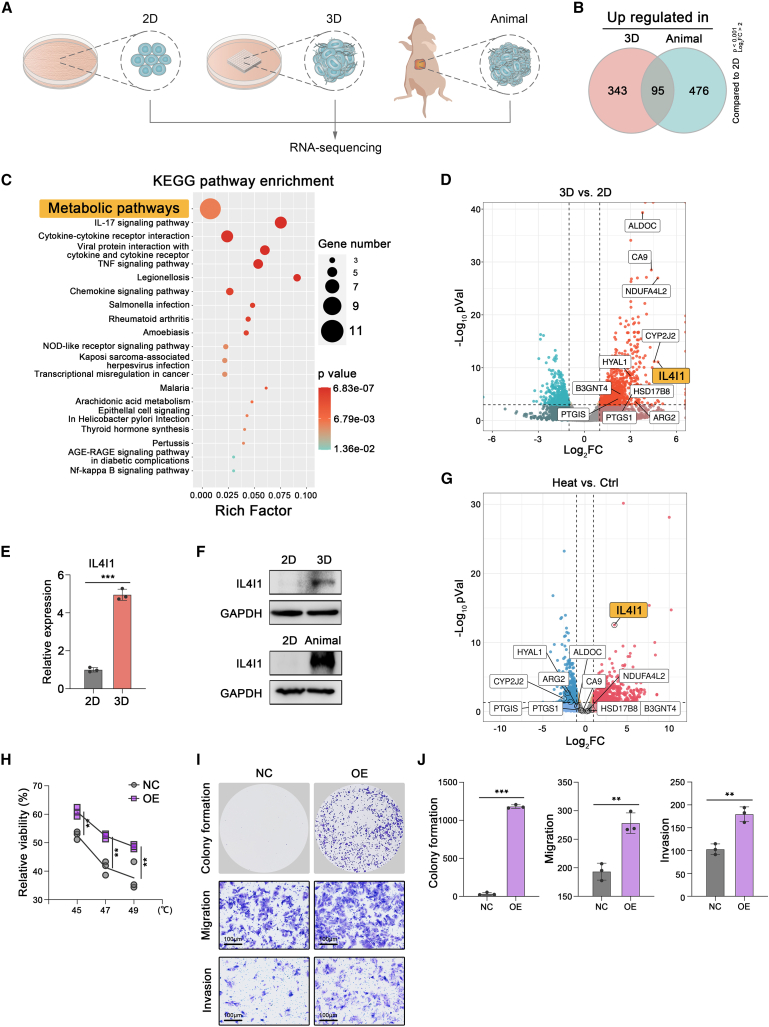
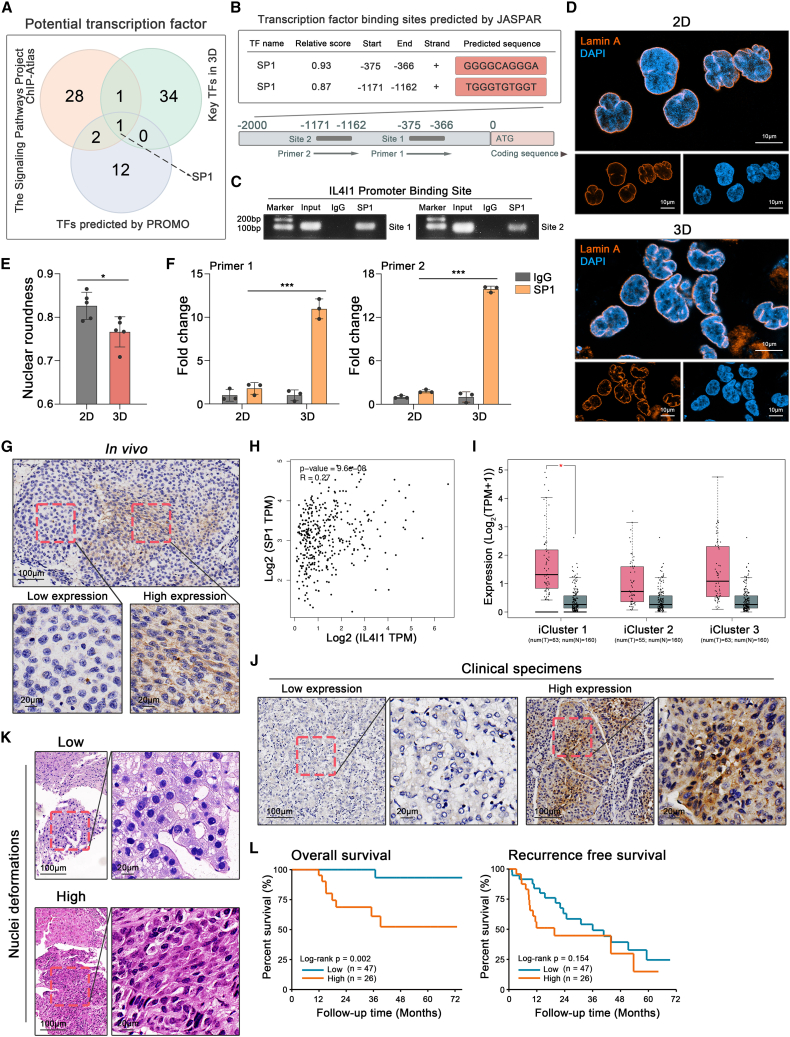
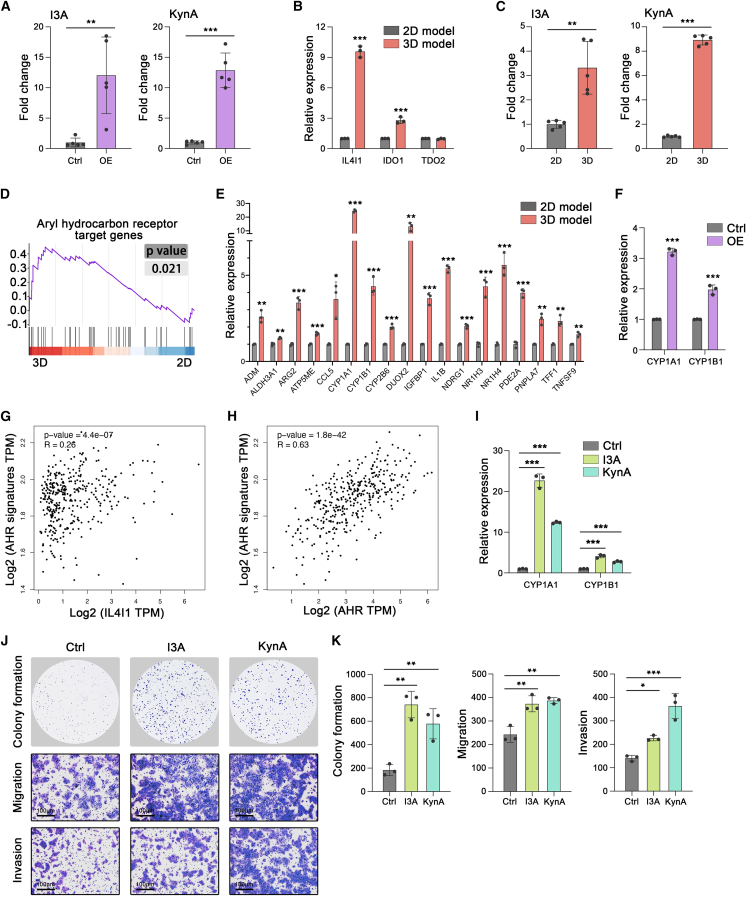
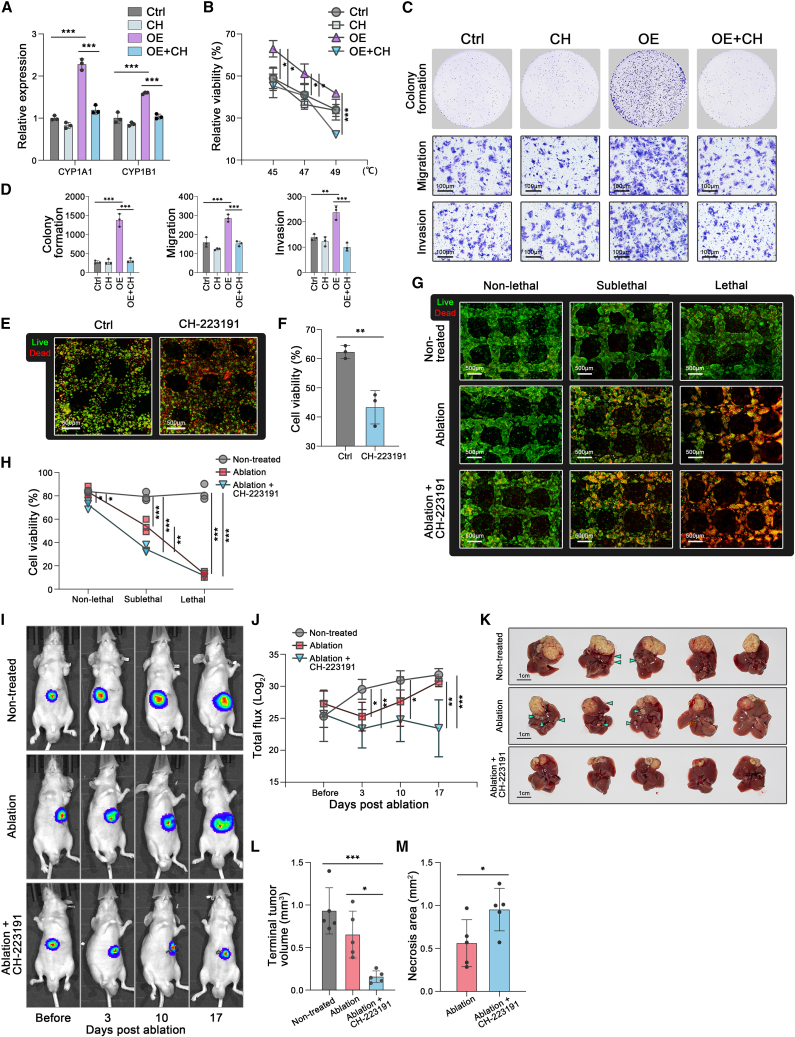
Mechanical stress can modulate the fate of cells in both physiological and extreme conditions. Recurrence of tumors after thermal ablation, a radical therapy for many cancers, indicates that some tumor cells can endure temperatures far beyond physiological ones. This unusual heat resistance with unknown mechanisms remains a key obstacle to fully realizing the clinical potential of thermal ablation. By developing a 3D bioprinting-based thermal ablation system, we demonstrate that hepatocellular carcinoma (HCC) cells in this 3D model exhibit enhanced heat resistance as compared with cells on plates. Mechanistically, the activation of transcription factor SP1 under mechanical confinement enhances the transcription of Interleukin-4-Induced-1, which catalyzes tryptophan metabolites to activate the aryl hydrocarbon receptor (AHR), leading to heat resistance. Encouragingly, the AHR inhibitor prevents HCC recurrence after thermal ablation. These findings reveal a previously unknown role of mechanical confinement in heat resistance and provide a rationale for AHR inhibitors as neoadjuvant therapy.
Keywords: heat resistance; hepatocellular carcinoma; mechanical confinement; nucleus deformations; tryptophan metabolism.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Aryl hydrocarbon receptor promotes hepatocellular carcinoma tumorigenesis by targeting intestine-specific homeobox expression.Mol Carcinog. 2017 Oct;56(10):2167-2177. doi: 10.1002/mc.22658. Epub 2017 Jul 28. Mol Carcinog. 2017. PMID: 28398627
-
IL4I1 Is a Metabolic Immune Checkpoint that Activates the AHR and Promotes Tumor Progression.Cell. 2020 Sep 3;182(5):1252-1270.e34. doi: 10.1016/j.cell.2020.07.038. Epub 2020 Aug 19. Cell. 2020. PMID: 32818467
-
NUPR1 contributes to radiation resistance by maintaining ROS homeostasis via AhR/CYP signal axis in hepatocellular carcinoma.BMC Med. 2022 Oct 19;20(1):365. doi: 10.1186/s12916-022-02554-3. BMC Med. 2022. PMID: 36258210 Free PMC article.
-
Host/microbiota interactions-derived tryptophan metabolites modulate oxidative stress and inflammation via aryl hydrocarbon receptor signaling.Free Radic Biol Med. 2022 May 1;184:30-41. doi: 10.1016/j.freeradbiomed.2022.03.025. Epub 2022 Mar 31. Free Radic Biol Med. 2022. PMID: 35367341 Review.
-
Nutritional Therapy to Modulate Tryptophan Metabolism and Aryl Hydrocarbon-Receptor Signaling Activation in Human Diseases.Nutrients. 2020 Sep 17;12(9):2846. doi: 10.3390/nu12092846. Nutrients. 2020. PMID: 32957545 Free PMC article. Review.
Cited by
-
Collagen extracellular matrix promotes gastric cancer immune evasion by activating IL4I1-AHR signaling.Transl Oncol. 2024 Nov;49:102113. doi: 10.1016/j.tranon.2024.102113. Epub 2024 Aug 30. Transl Oncol. 2024. PMID: 39216468 Free PMC article.
-
IL4I1 in M2-like macrophage promotes glioma progression and is a promising target for immunotherapy.Front Immunol. 2024 Jan 5;14:1338244. doi: 10.3389/fimmu.2023.1338244. eCollection 2023. Front Immunol. 2024. PMID: 38250074 Free PMC article.
-
Regulating macrophage phenotypes with IL4I1-mimetic nanoparticles in IDD treatment.J Nanobiotechnology. 2025 Mar 6;23(1):175. doi: 10.1186/s12951-025-03241-0. J Nanobiotechnology. 2025. PMID: 40050923 Free PMC article.
-
Nanostrategies synergize with locoregional interventional therapies for boosting antitumor immunity.Bioact Mater. 2025 May 31;51:634-649. doi: 10.1016/j.bioactmat.2025.05.016. eCollection 2025 Sep. Bioact Mater. 2025. PMID: 40521175 Free PMC article.
-
PM2.5 Induced Nasal Mucosal Barrier Dysfunction and Epithelial-Mesenchymal Transition to Promote Chronic Rhinosinusitis Through IL4I1-AhR Signaling Pathway.Toxics. 2025 Jun 10;13(6):488. doi: 10.3390/toxics13060488. Toxics. 2025. PMID: 40559961 Free PMC article.
References
-
- Nader G.P.d.F., Agüera-Gonzalez S., Routet F., Gratia M., Maurin M., Cancila V., Cadart C., Palamidessi A., Ramos R.N., San Roman M., et al. Compromised nuclear envelope integrity drives TREX1-dependent DNA damage and tumor cell invasion. Cell. 2021;184:5230–5246.e22. doi: 10.1016/j.cell.2021.08.035. - DOI - PubMed
-
- Stowers R.S., Shcherbina A., Israeli J., Gruber J.J., Chang J., Nam S., Rabiee A., Teruel M.N., Snyder M.P., Kundaje A., Chaudhuri O. Matrix stiffness induces a tumorigenic phenotype in mammary epithelium through changes in chromatin accessibility. Nat. Biomed. Eng. 2019;3:1009–1019. doi: 10.1038/s41551-019-0420-5. - DOI - PMC - PubMed
-
- Nava M.M., Miroshnikova Y.A., Biggs L.C., Whitefield D.B., Metge F., Boucas J., Vihinen H., Jokitalo E., Li X., García Arcos J.M., et al. Heterochromatin-Driven Nuclear Softening Protects the Genome against Mechanical Stress-Induced Damage. Cell. 2020;181:800–817.e22. doi: 10.1016/j.cell.2020.03.052. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
