

Premature aging and reduced cancer incidence associated with near-complete body-wide Myc inactivation
- PMID: 37481724
- PMCID: PMC10591215
- DOI: 10.1016/j.celrep.2023.112830
Premature aging and reduced cancer incidence associated with near-complete body-wide Myc inactivation
Abstract
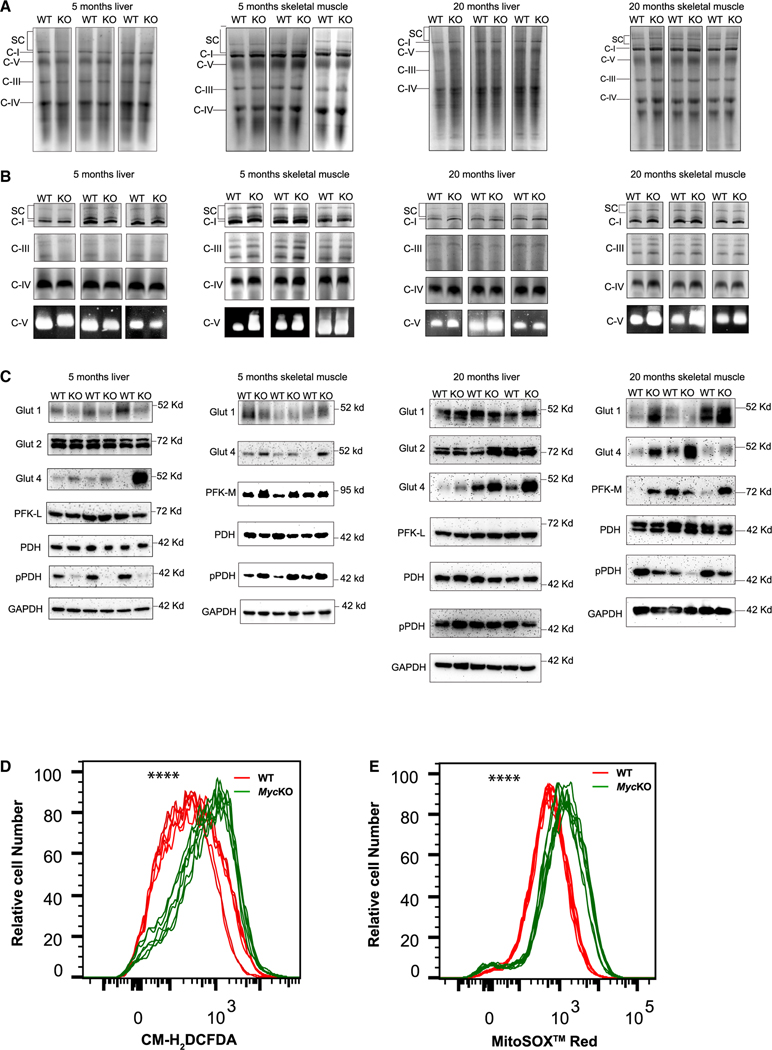
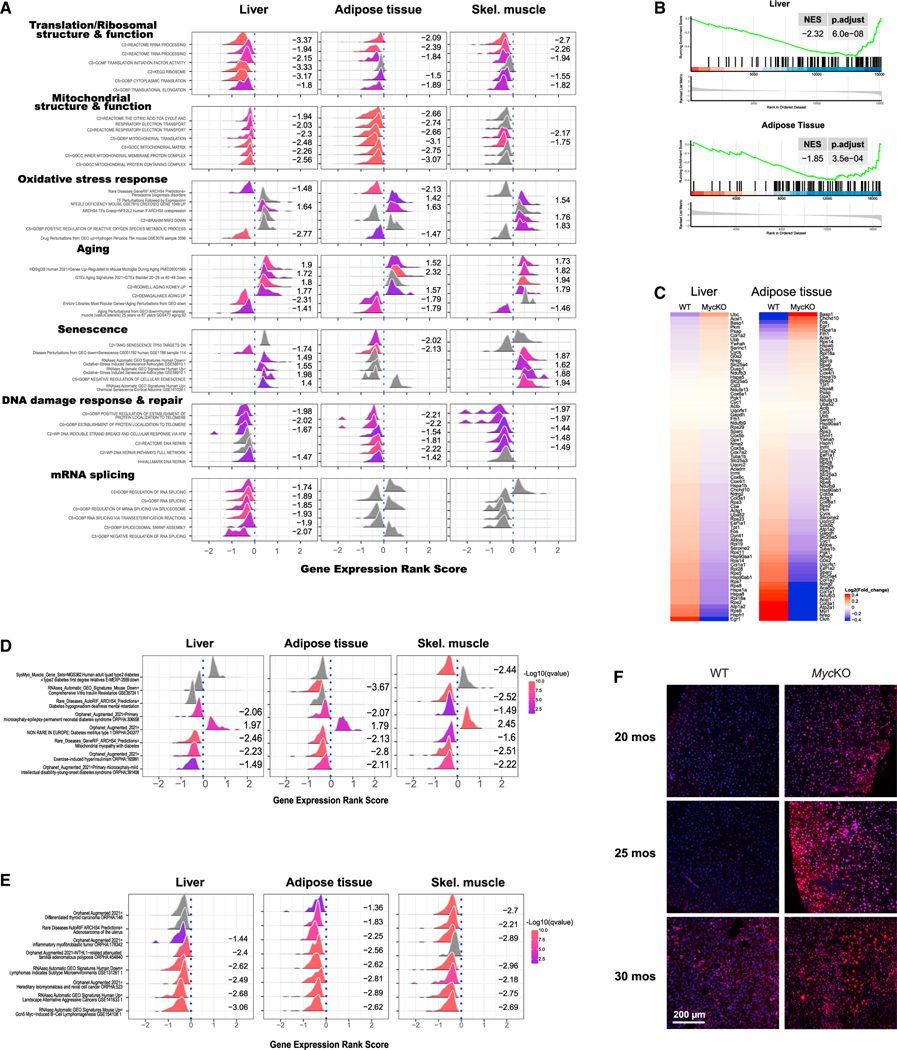
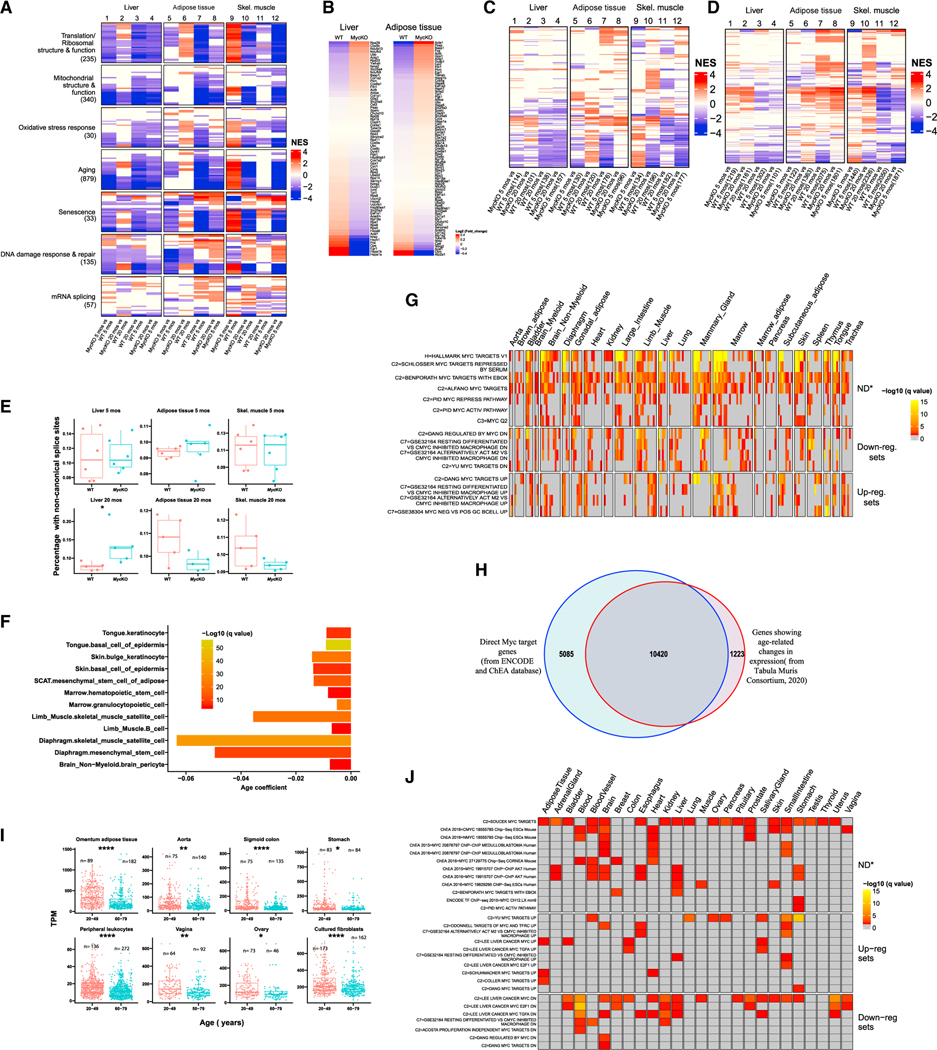
MYC proto-oncogene dysregulation alters metabolism, translation, and other functions in ways that support tumor induction and maintenance. Although Myc+/- mice are healthier and longer-lived than control mice, the long-term ramifications of more complete Myc loss remain unknown. We now describe the chronic consequences of body-wide Myc inactivation initiated postnatally. "MycKO" mice acquire numerous features of premature aging, including altered body composition and habitus, metabolic dysfunction, hepatic steatosis, and dysregulation of gene sets involved in functions that normally deteriorate with aging. Yet, MycKO mice have extended lifespans that correlate with a 3- to 4-fold lower lifetime cancer incidence. Aging tissues from normal mice and humans also downregulate Myc and gradually alter many of the same Myc target gene sets seen in MycKO mice. Normal aging and its associated cancer predisposition are thus highly linked via Myc.
Keywords: CP: Cancer; aging.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
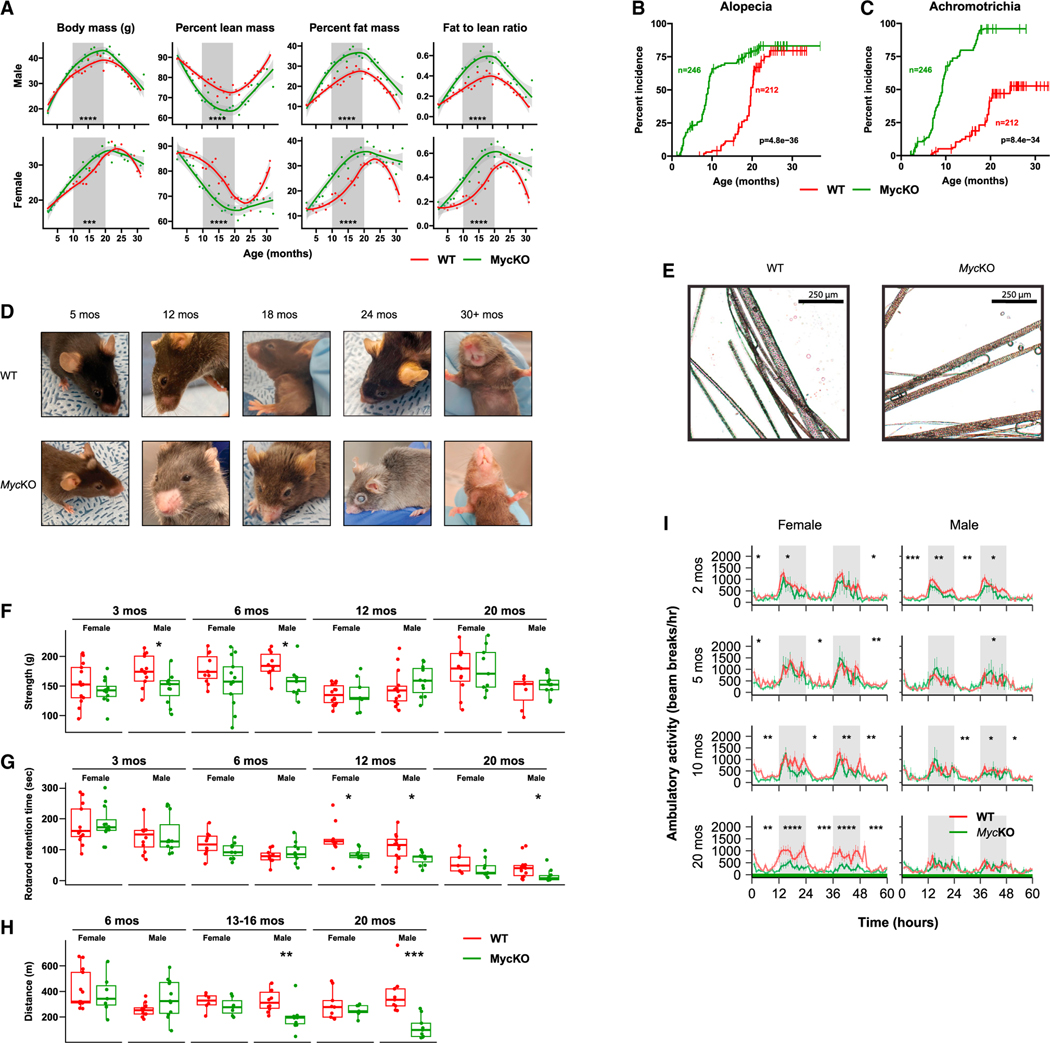
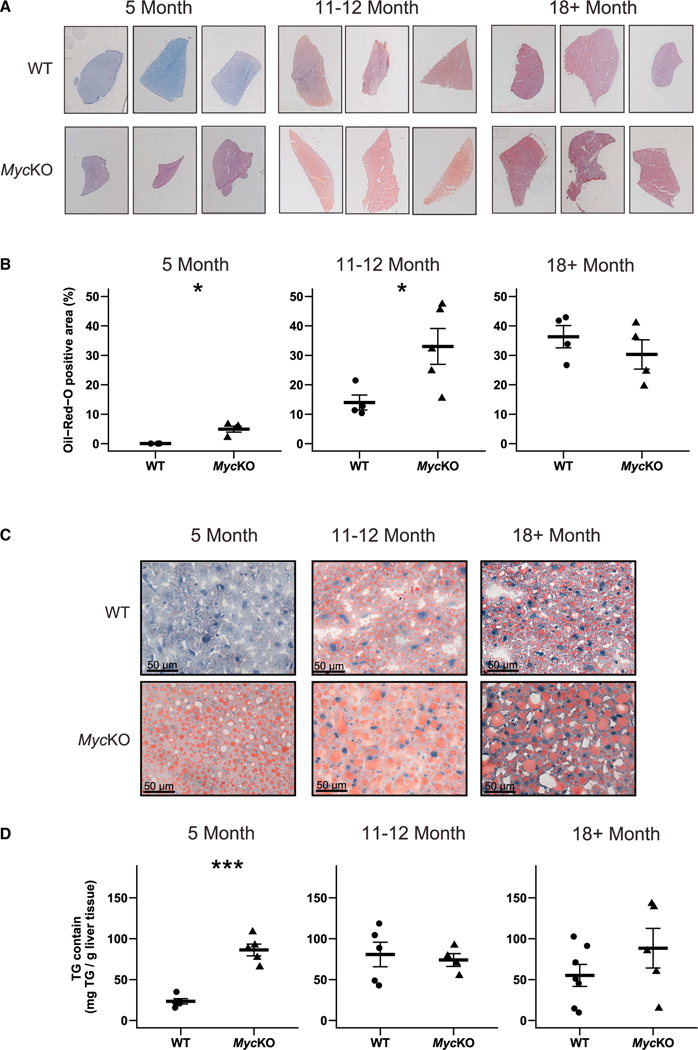
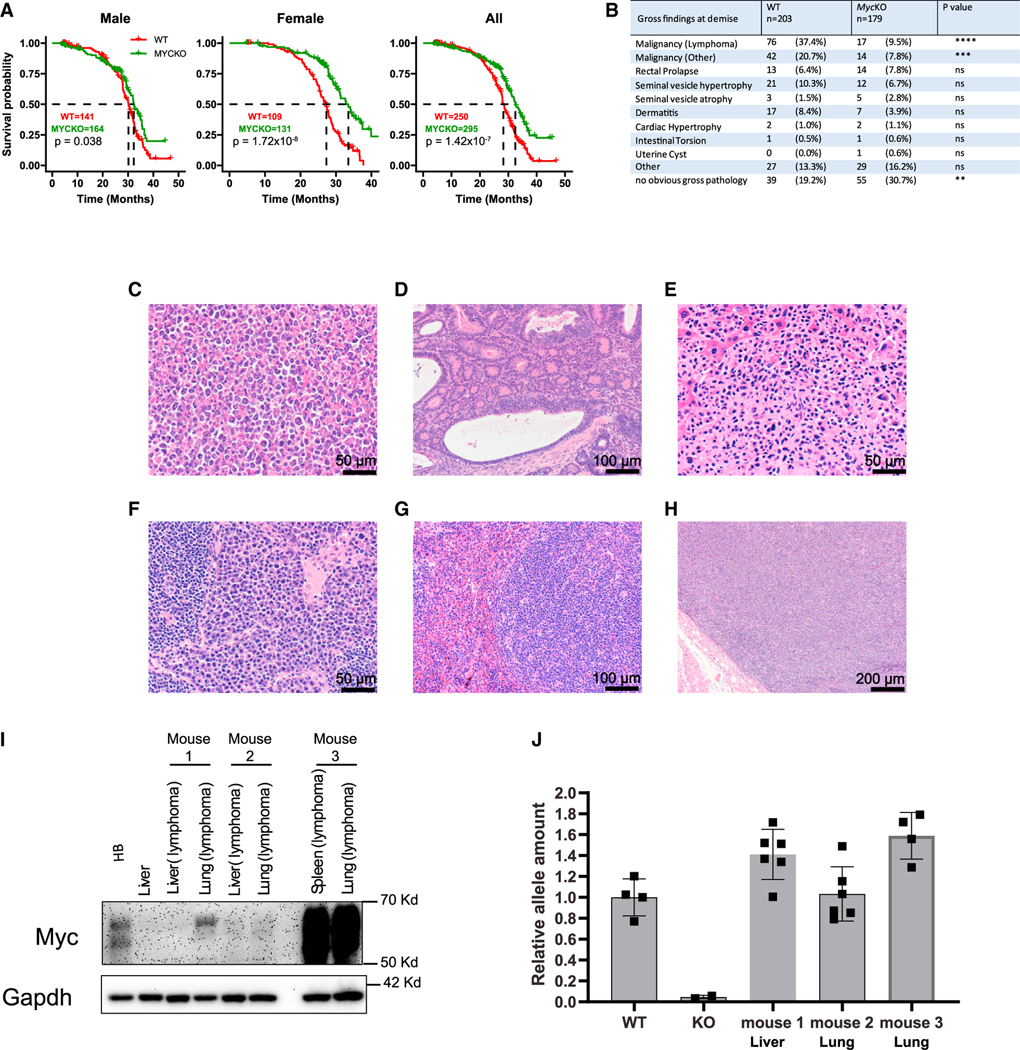
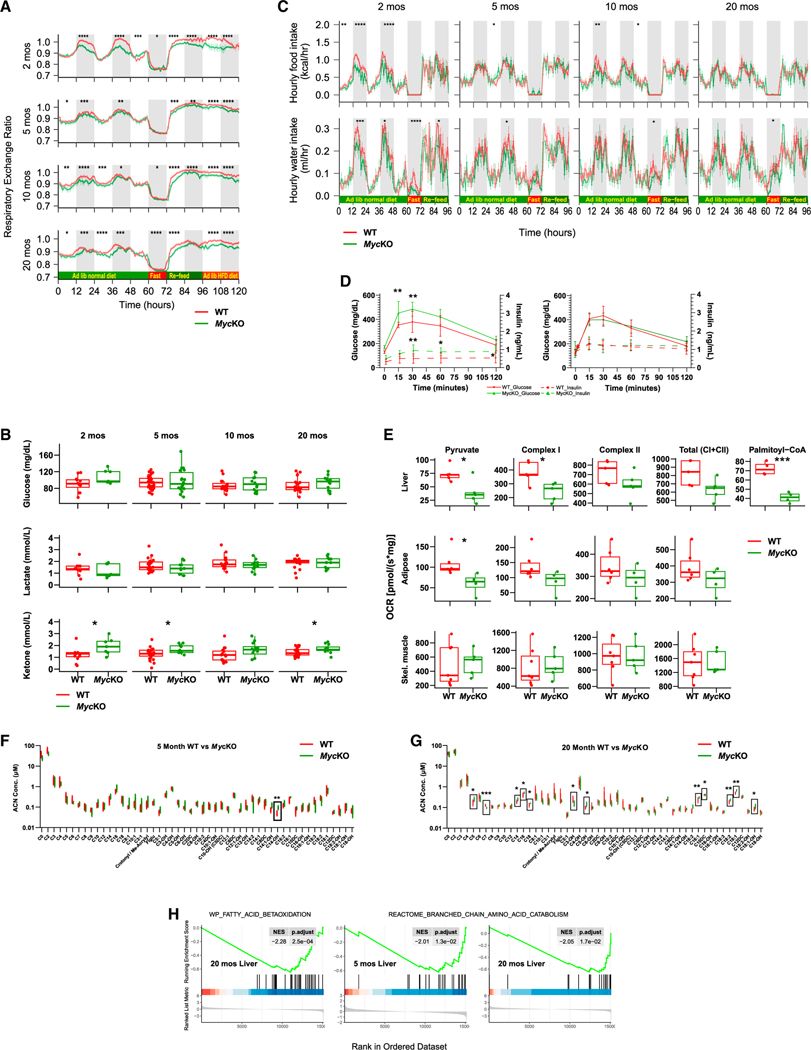
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
