

The mechanical stability of proteins regulates their translocation rate into the cell nucleus
- PMID: 37484710
- PMCID: PMC7614795
- DOI: 10.1038/s41567-019-0551-3
The mechanical stability of proteins regulates their translocation rate into the cell nucleus
Abstract
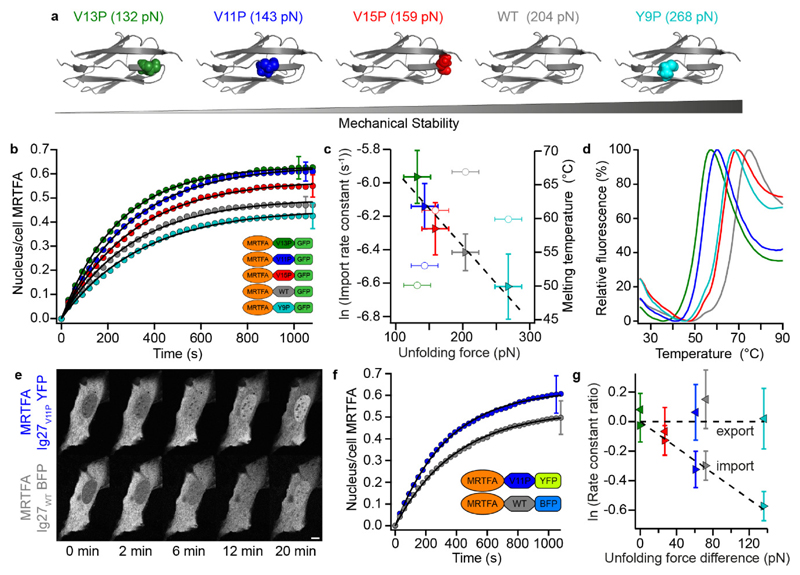
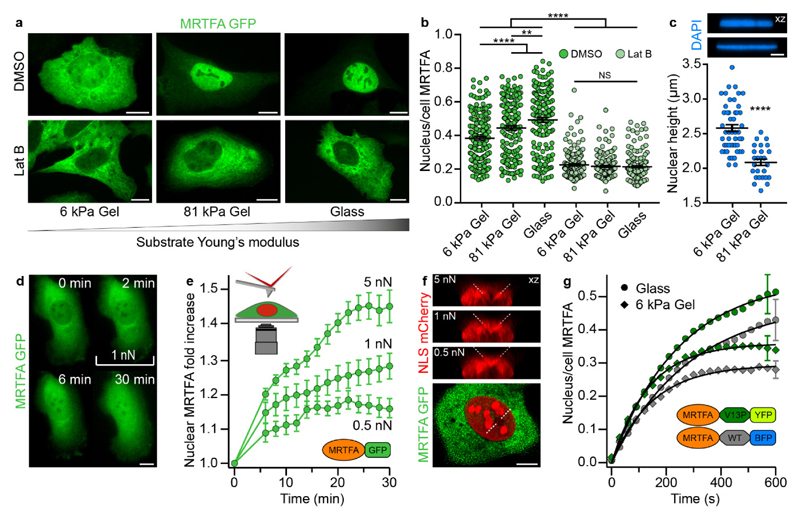
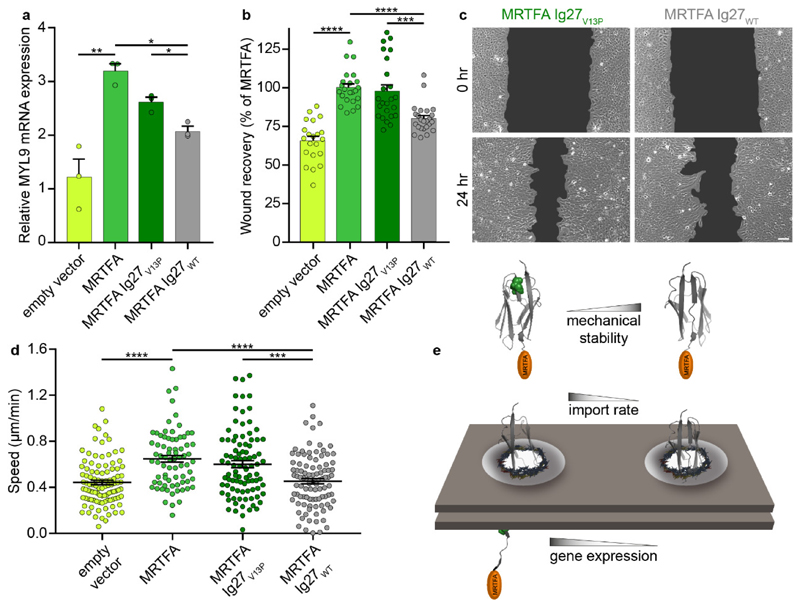
The translocation of mechanosensitive transcription factors (TFs) across the nuclear envelope is a crucial step in cellular mechanotransduction. Yet the molecular mechanisms by which external mechanical cues control the nuclear shuttling dynamics of TFs through the nuclear pore complex (NPC) to activate gene expression are poorly understood. Here, we show that the nuclear import rate of myocardin-related transcription factor A (MRTFA) - a protein that regulates cytoskeletal dynamics via the activation of the TF serum response factor (SRF) - inversely correlates with the protein's nanomechanical stability and does not relate to its thermodynamic stability. Tagging MRTFA with mechanically resistant proteins results in the downregulation of SRF-mediated myosin light-chain 9 (MYL9) gene expression and subsequent slowing down of cell migration. We conclude that the mechanical unfolding of proteins regulates their nuclear translocation rate through the NPC, and highlight the role of the NPC as a selective mechanosensor able to discriminate forces as low as ~10 pN. The modulation of the mechanical stability of TFs may represent a new strategy for the control of gene expression.
Conflict of interest statement
Competing financial interests: The authors declare no competing financial interests.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous