

Molecular signatures define subtypes of auditory afferents with distinct peripheral projection patterns and physiological properties
- PMID: 37487063
- PMCID: PMC10400978
- DOI: 10.1073/pnas.2217033120
Molecular signatures define subtypes of auditory afferents with distinct peripheral projection patterns and physiological properties
Abstract
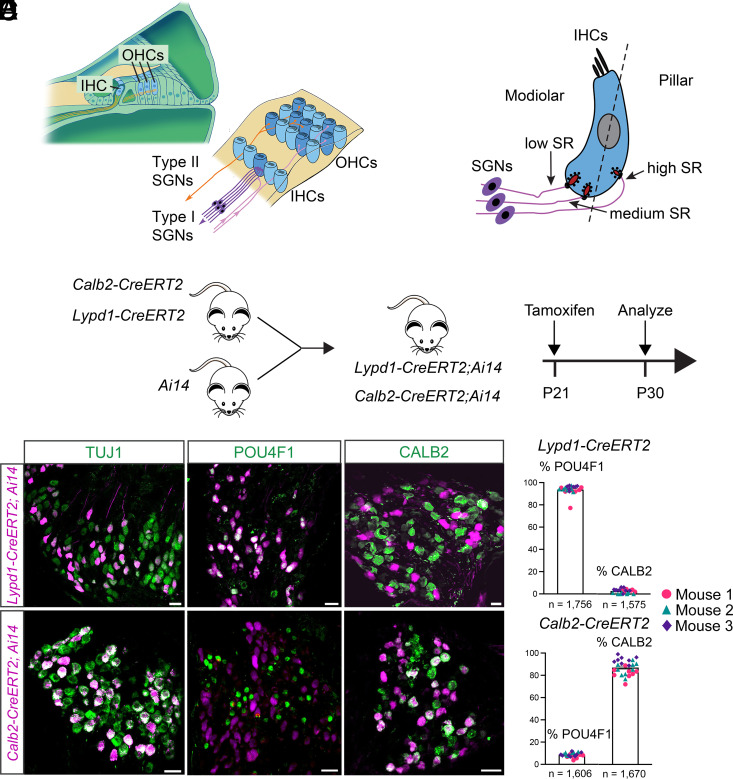
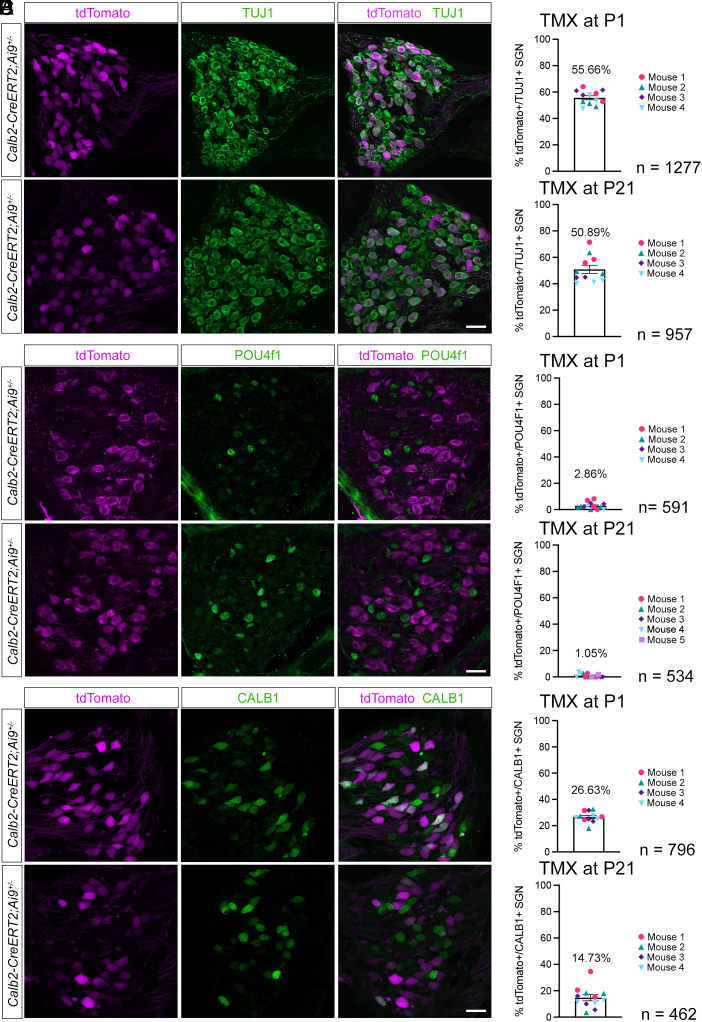
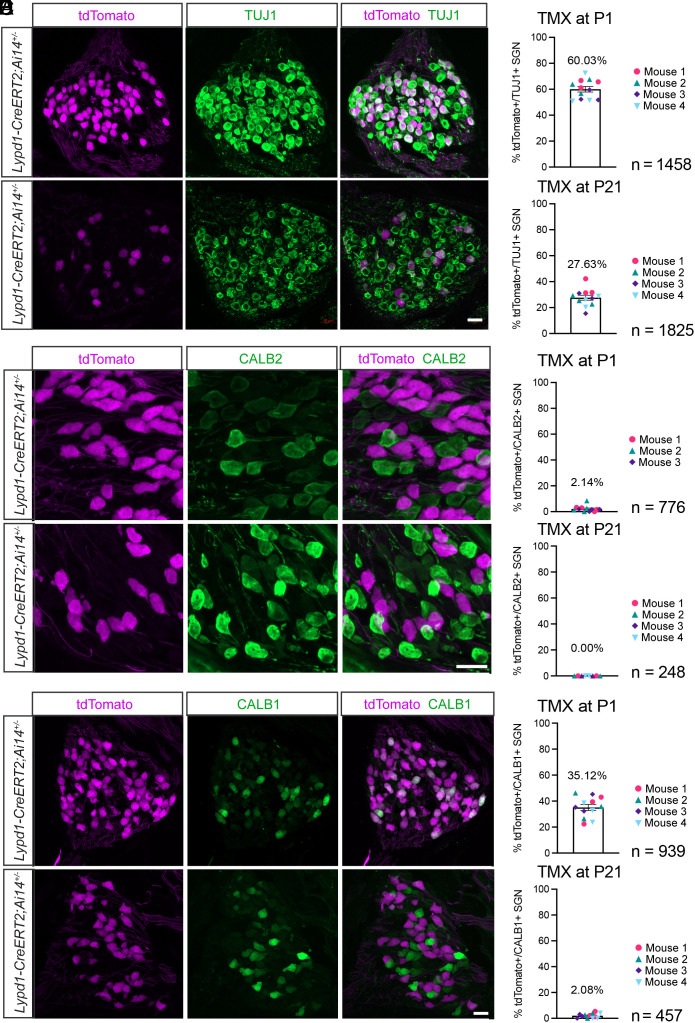
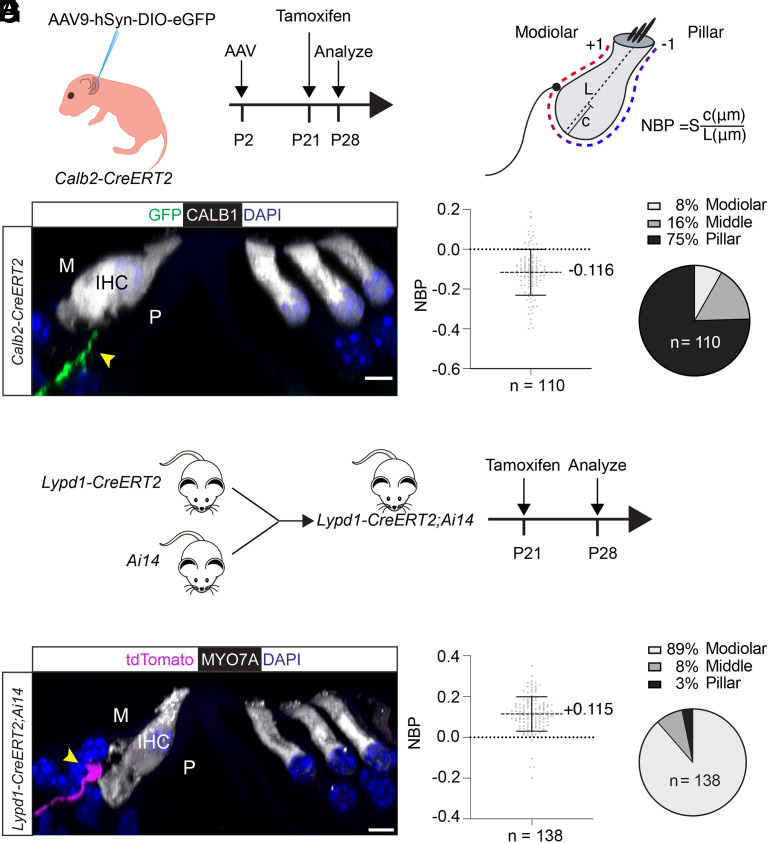
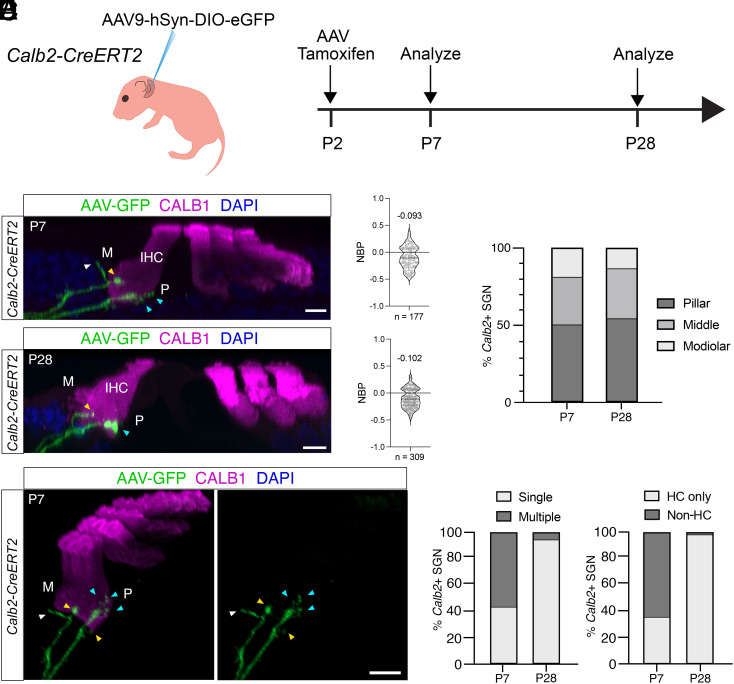
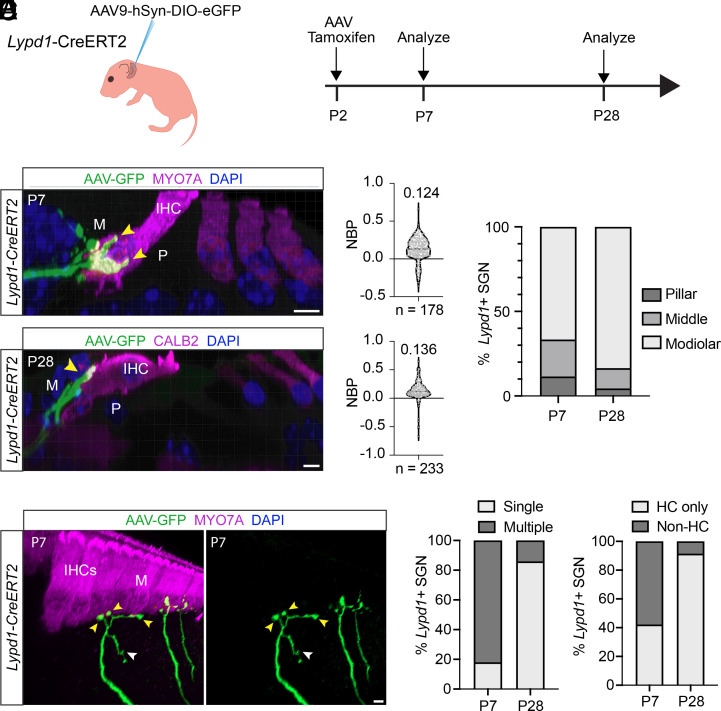
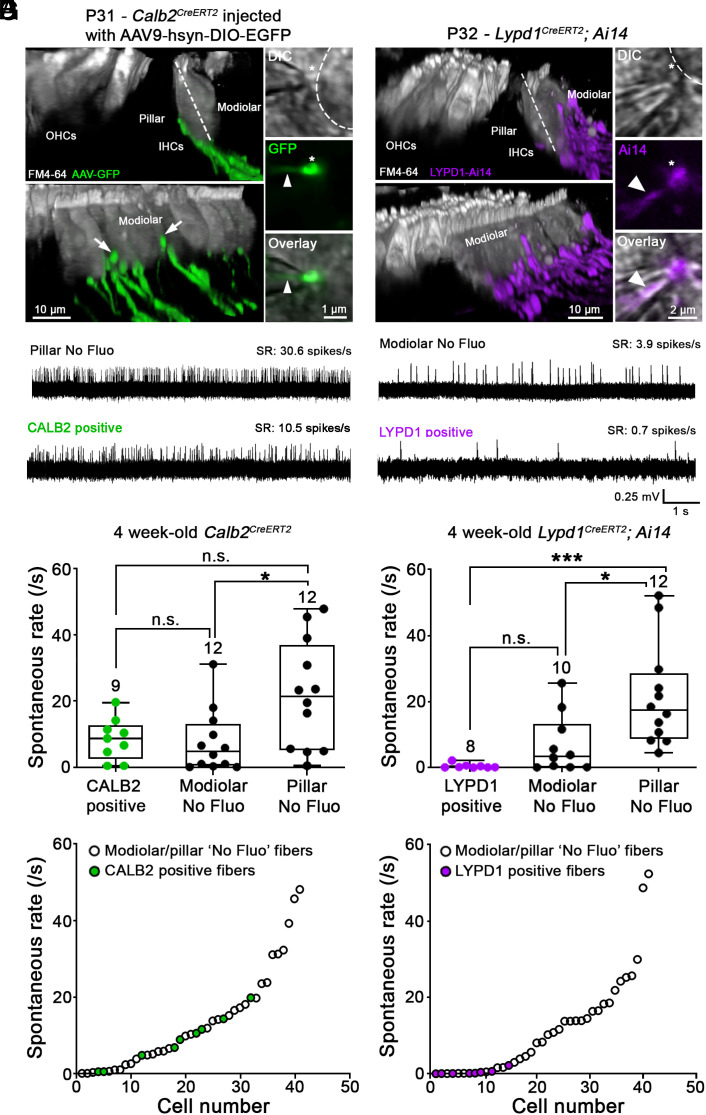
Type I spiral ganglion neurons (SGNs) are the auditory afferents that transmit sound information from cochlear inner hair cells (IHCs) to the brainstem. These afferents consist of physiological subtypes that differ in their spontaneous firing rate (SR), activation threshold, and dynamic range and have been described as low, medium, and high SR fibers. Lately, single-cell RNA sequencing experiments have revealed three molecularly defined type I SGN subtypes. The extent to which physiological type I SGN subtypes correspond to molecularly defined subtypes is unclear. To address this question, we have generated mouse lines expressing CreERT2 in SGN subtypes that allow for a physiological assessment of molecular subtypes. We show that Lypd1-CreERT2 expressing SGNs represent a well-defined group of neurons that preferentially innervate the IHC modiolar side and exhibit a narrow range of low SRs. In contrast, Calb2-CreERT2 expressing SGNs preferentially innervate the IHC pillar side and exhibit a wider range of SRs, thus suggesting that a strict stratification of all SGNs into three molecular subclasses is not obvious, at least not with the CreERT2 tools used here. Genetically marked neuronal subtypes refine their innervation specificity onto IHCs postnatally during the time when activity is required to refine their molecular phenotype. Type I SGNs thus consist of genetically defined subtypes with distinct physiological properties and innervation patterns. The molecular subtype-specific lines characterized here will provide important tools for investigating the role of the physiologically distinct type I SGNs in encoding sound signals.
Keywords: CreERT2; hair cell; hearing; inner ear; spiral ganglion neuron.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Sanes J. R., Masland R. H., The types of retinal ganglion cells: Current status and implications for neuronal classification. Annu. Rev. Neurosci. 38, 221–246 (2015). - PubMed
-
- Seabrook T. A., Burbridge T. J., Crair M. C., Huberman A. D., Architecture, function, and assembly of the mouse visual system. Annu. Rev. Neurosci. 40, 499–538 (2017). - PubMed
-
- Heil P., Peterson A. J., Basic response properties of auditory nerve fibers: A review. Cell Tissue Res. 361, 129–158 (2015). - PubMed
-
- Oxenham A. J., Plack C. J., A behavioral measure of basilar-membrane nonlinearity in listeners with normal and impaired hearing. J. Acoust. Soc. Am. 101, 3666–3675 (1997). - PubMed
-
- Reijntjes D. O. J., Pyott S. J., The afferent signaling complex: Regulation of type I spiral ganglion neuron responses in the auditory periphery. Hear Res. 336, 1–16 (2016). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
