

Chromatin architectural factor CTCF is essential for progesterone-dependent uterine maturation
- PMID: 37489832
- PMCID: PMC10372848
- DOI: 10.1096/fj.202300862R
Chromatin architectural factor CTCF is essential for progesterone-dependent uterine maturation
Abstract
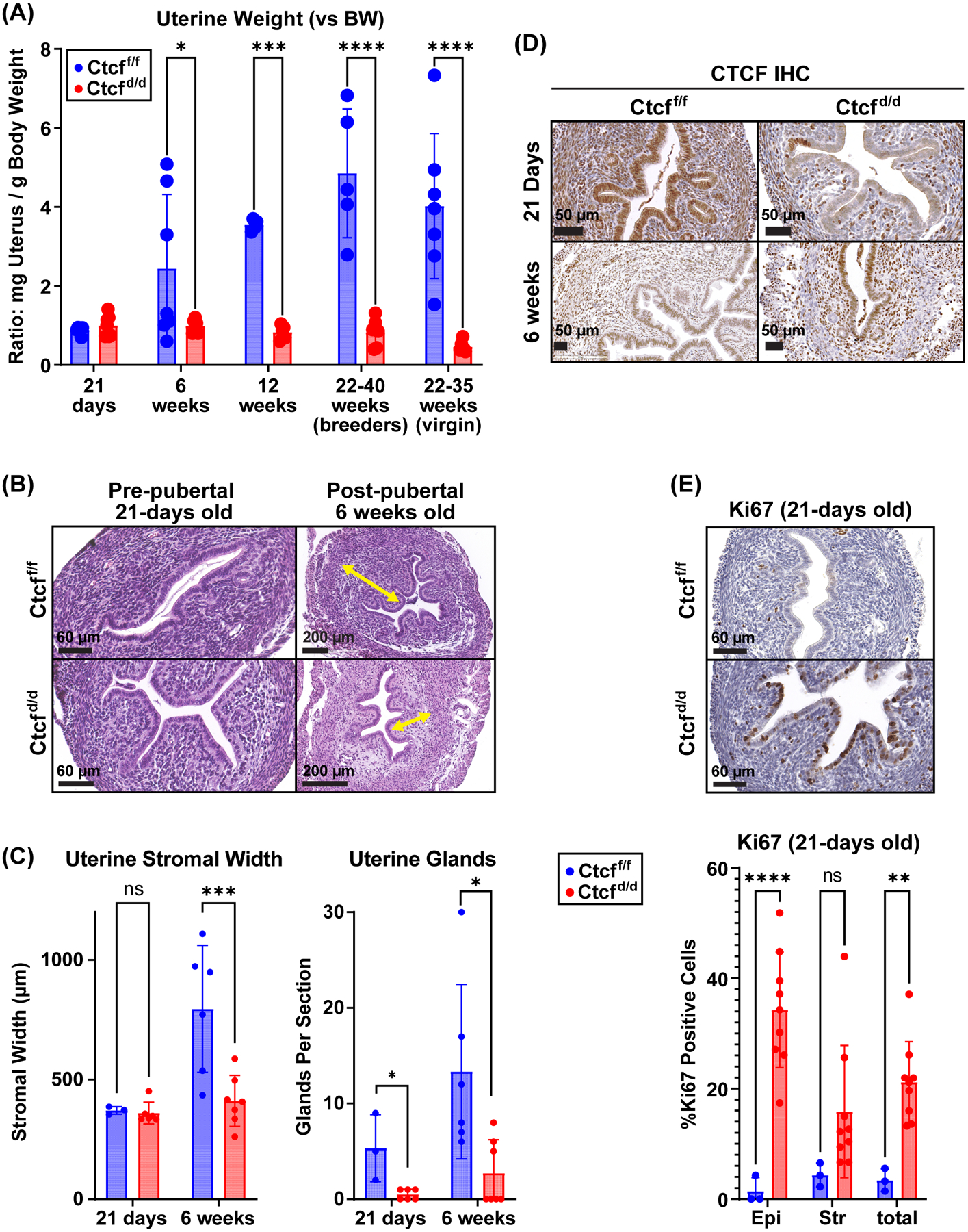
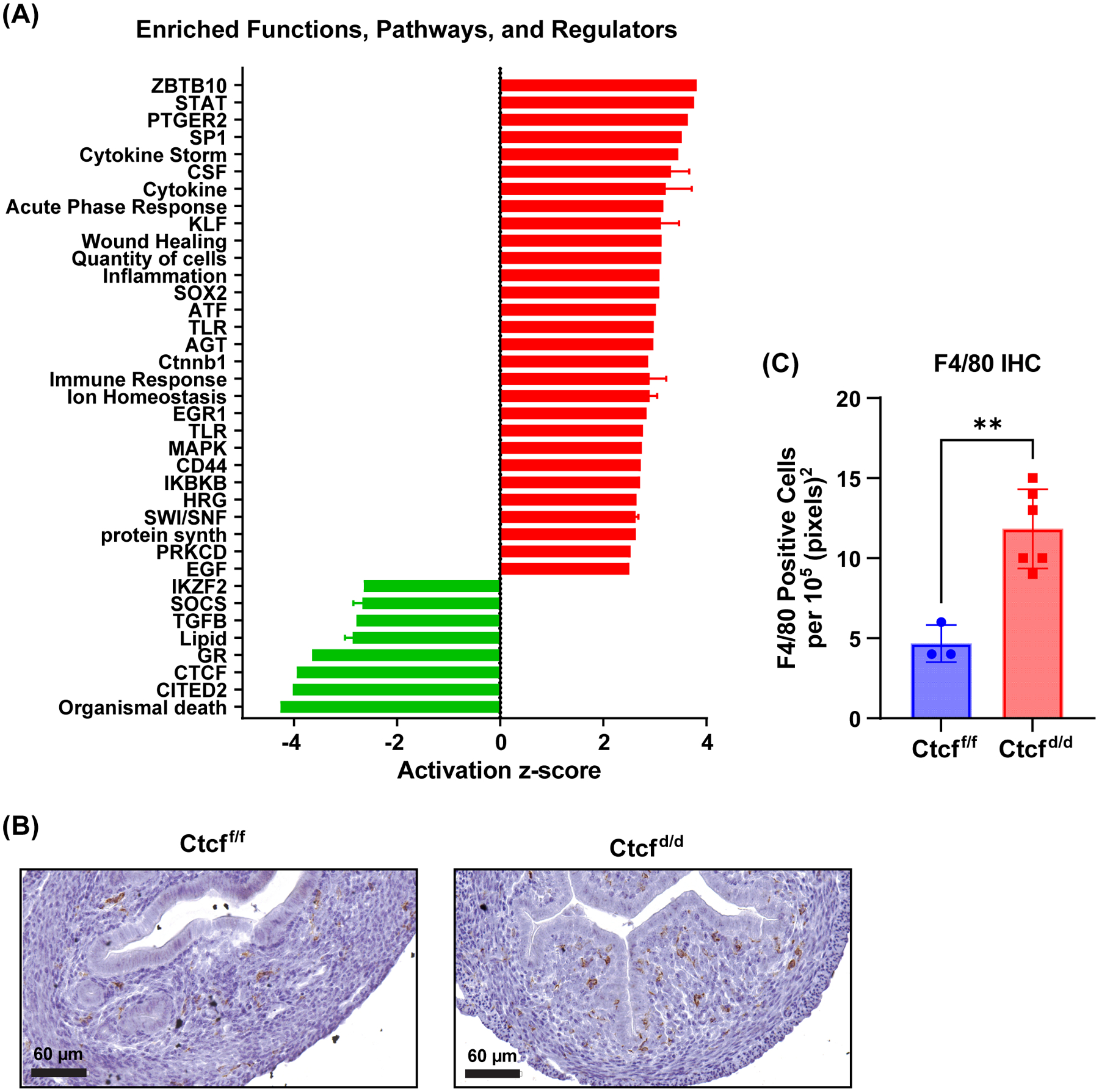
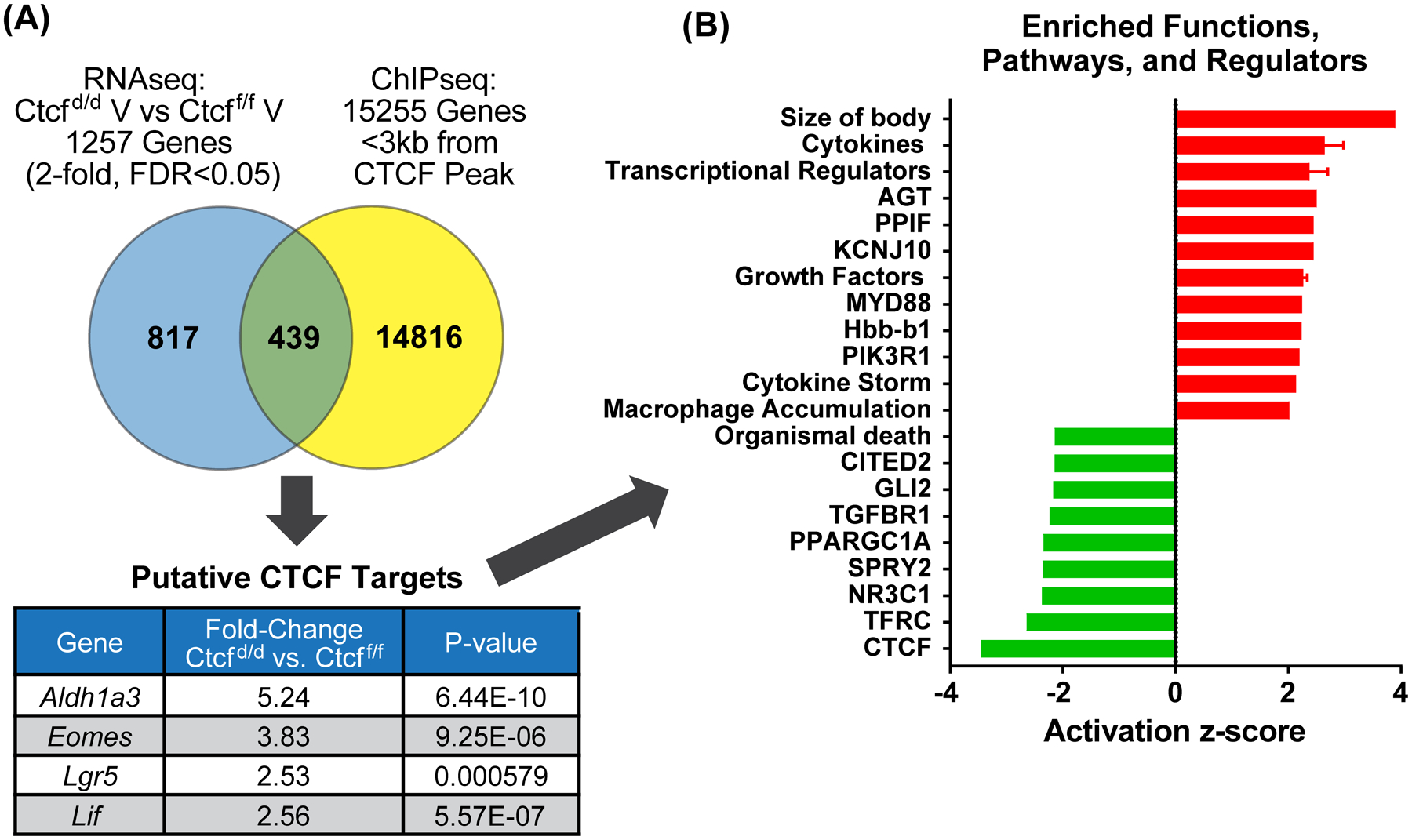
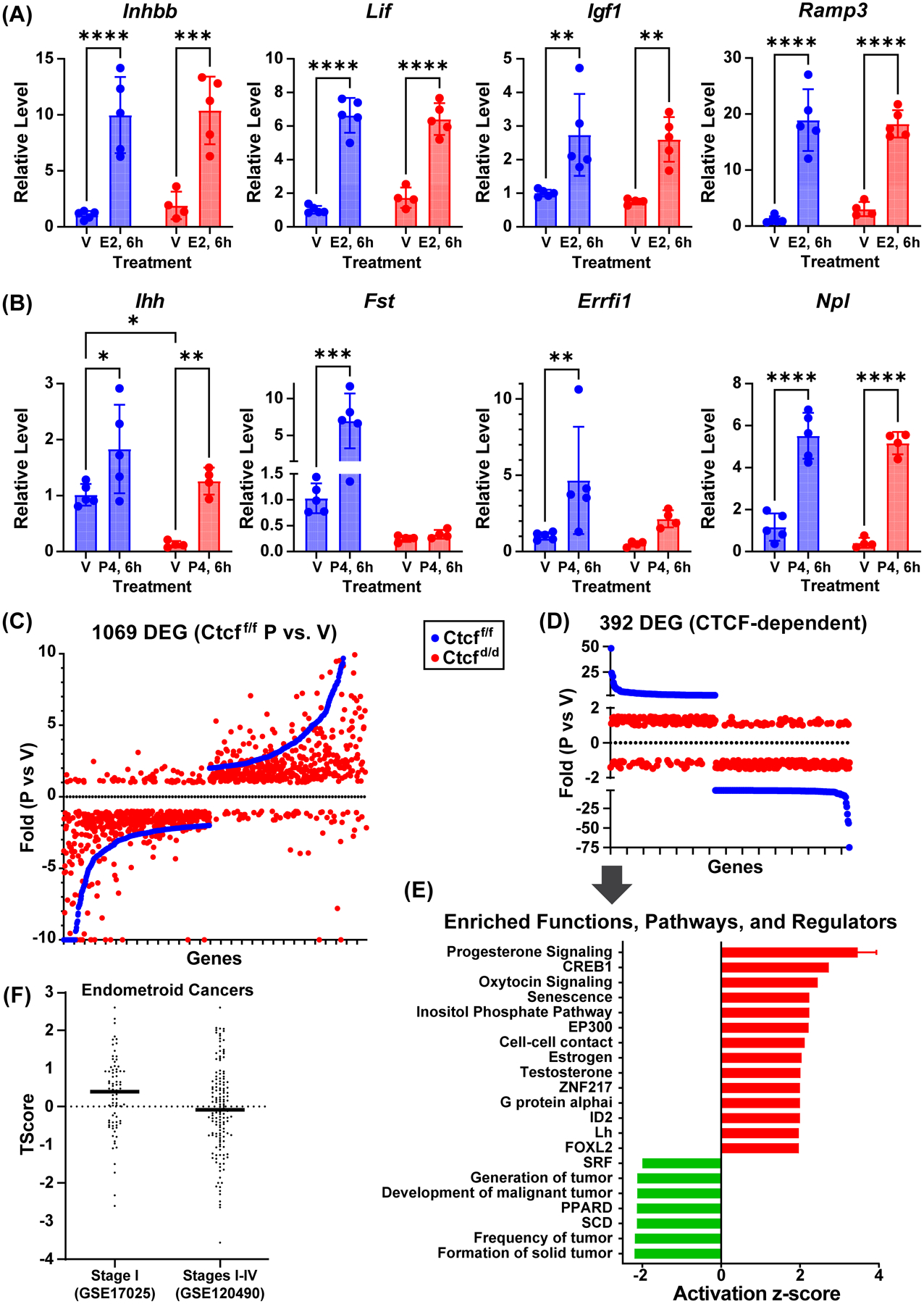
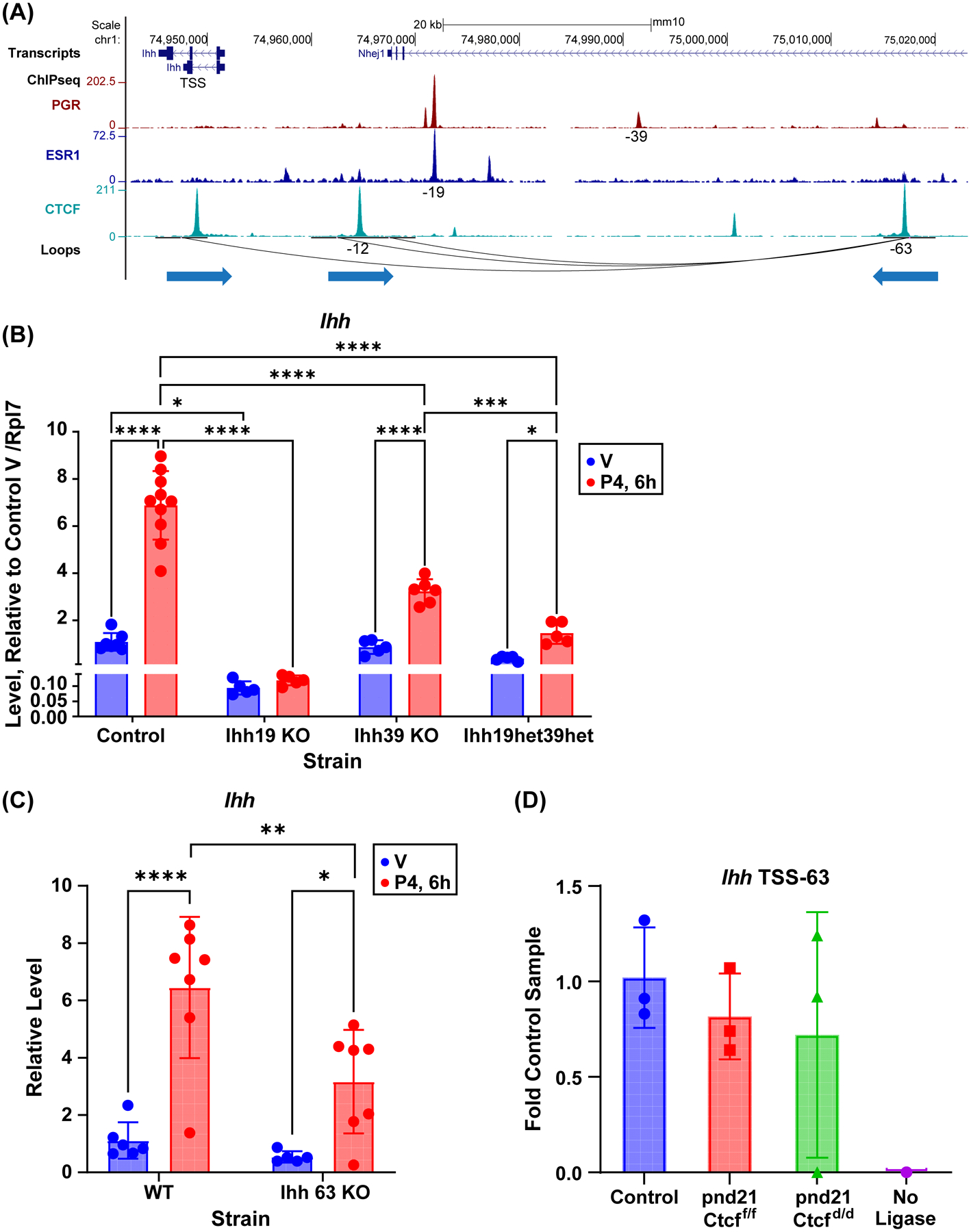
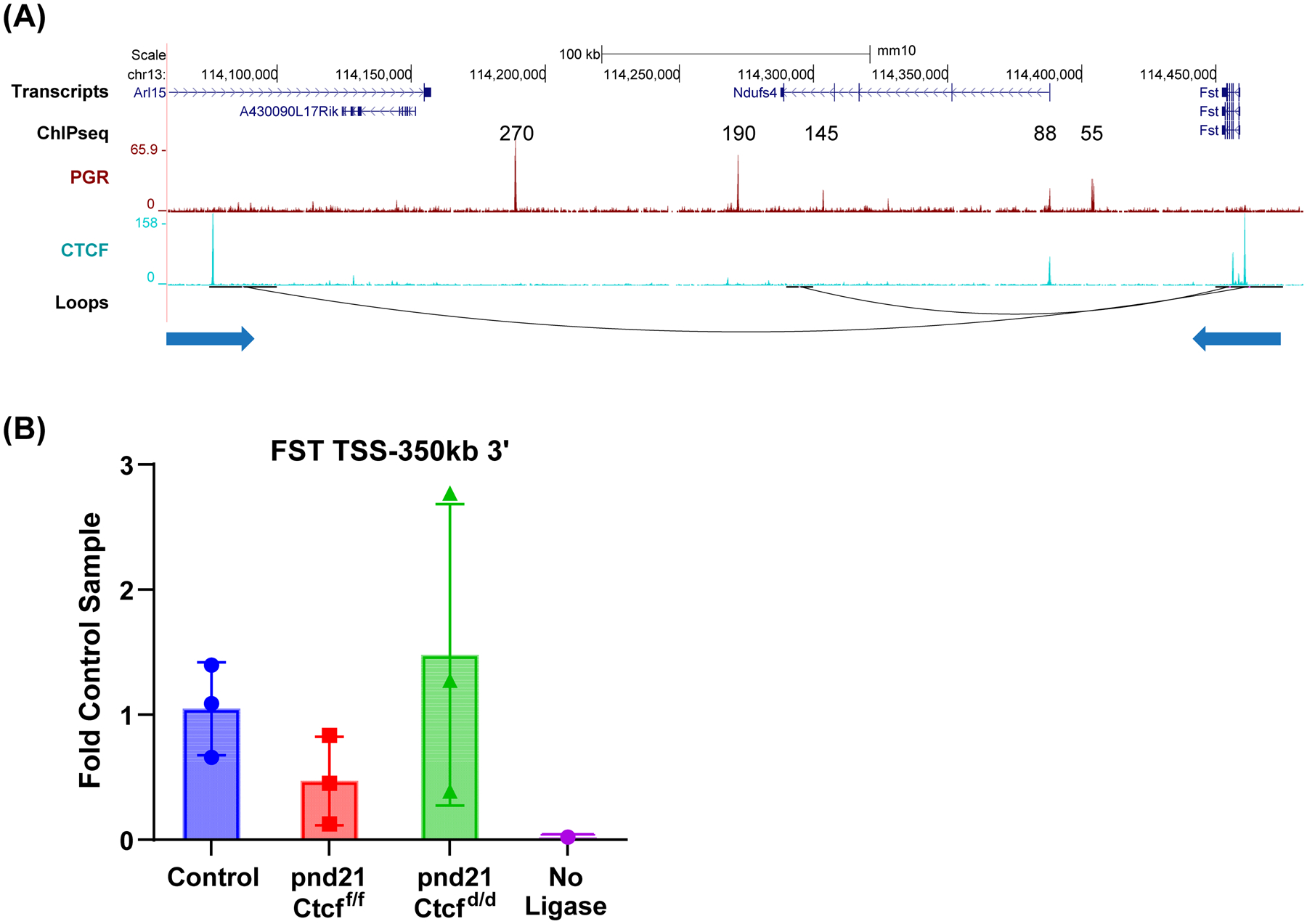
Receptors for estrogen and progesterone frequently interact, via Cohesin/CTCF loop extrusion, at enhancers distal from regulated genes. Loss-of-function CTCF mutation in >20% of human endometrial tumors indicates its importance in uterine homeostasis. To better understand how CTCF-mediated enhancer-gene interactions impact endometrial development and function, the Ctcf gene was selectively deleted in female reproductive tissues of mice. Prepubertal Ctcfd/d uterine tissue exhibited a marked reduction in the number of uterine glands compared to those without Ctcf deletion (Ctcff/f mice). Post-pubertal Ctcfd/d uteri were hypoplastic with significant reduction in both the amount of the endometrial stroma and number of glands. Transcriptional profiling revealed increased expression of stem cell molecules Lif, EOMES, and Lgr5, and enhanced inflammation pathways following Ctcf deletion. Analysis of the response of the uterus to steroid hormone stimulation showed that CTCF deletion affects a subset of progesterone-responsive genes. This finding indicates (1) Progesterone-mediated signaling remains functional following Ctcf deletion and (2) certain progesterone-regulated genes are sensitive to Ctcf deletion, suggesting they depend on gene-enhancer interactions that require CTCF. The progesterone-responsive genes altered by CTCF ablation included Ihh, Fst, and Errfi1. CTCF-dependent progesterone-responsive uterine genes enhance critical processes including anti-tumorigenesis, which is relevant to the known effectiveness of progesterone in inhibiting progression of early-stage endometrial tumors. Overall, our findings reveal that uterine Ctcf plays a key role in progesterone-dependent expression of uterine genes underlying optimal post-pubertal uterine development.
Keywords: chromatin; estrogen; progesterone; puberty; uterus.
© 2023 Federation of American Societies for Experimental Biology. This article has been contributed to by U.S. Government employees and their work is in the public domain in the USA.
Conflict of interest statement
Conflict of Interest Statement
The authors declare no conflicts of interest
Figures
References
-
- Sikorska N, and Sexton T (2020) Defining Functionally Relevant Spatial Chromatin Domains: It is a TAD Complicated. Journal of Molecular Biology 432, 653–664 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
