

Killing in the name of: T6SS structure and effector diversity
- PMID: 37490402
- PMCID: PMC10433429
- DOI: 10.1099/mic.0.001367
Killing in the name of: T6SS structure and effector diversity
Abstract
The life of bacteria is challenging, to endure bacteria employ a range of mechanisms to optimize their environment, including deploying the type VI secretion system (T6SS). Acting as a bacterial crossbow, this system delivers effectors responsible for subverting host cells, killing competitors and facilitating general secretion to access common goods. Due to its importance, this lethal machine has been evolutionarily maintained, disseminated and specialized to fulfil these vital functions. In fact, T6SS structural clusters are present in over 25 % of Gram-negative bacteria, varying in number from one to six different genetic clusters per organism. Since its discovery in 2006, research on the T6SS has rapidly progressed, yielding remarkable breakthroughs. The identification and characterization of novel components of the T6SS, combined with biochemical and structural studies, have revealed fascinating mechanisms governing its assembly, loading, firing and disassembly processes. Recent findings have also demonstrated the efficacy of this system against fungal and Gram-positive cells, expanding its scope. Ongoing research continues to uncover an extensive and expanding repertoire of T6SS effectors, the genuine mediators of T6SS function. These studies are shedding light on new aspects of the biology of prokaryotic and eukaryotic organisms. This review provides a comprehensive overview of the T6SS, highlighting recent discoveries of its structure and the diversity of its effectors. Additionally, it injects a personal perspective on avenues for future research, aiming to deepen our understanding of this combative system.
Keywords: T6SS; T6SS assembly; Type VI secretion system; antieukaryotic effectors; antimicrobial effectors; baseplate; contractile sheath; inner tube; membrane complex; tail.
Conflict of interest statement
The authors declare no competing interest.
Figures
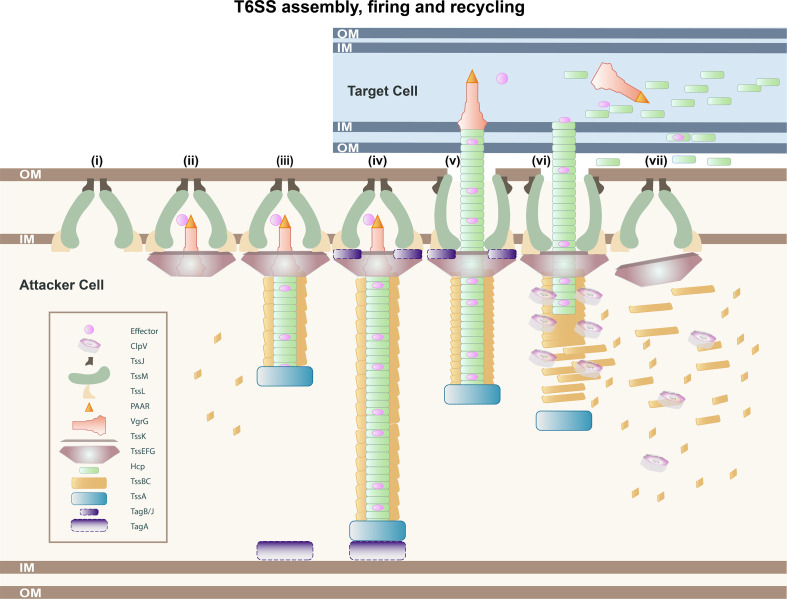
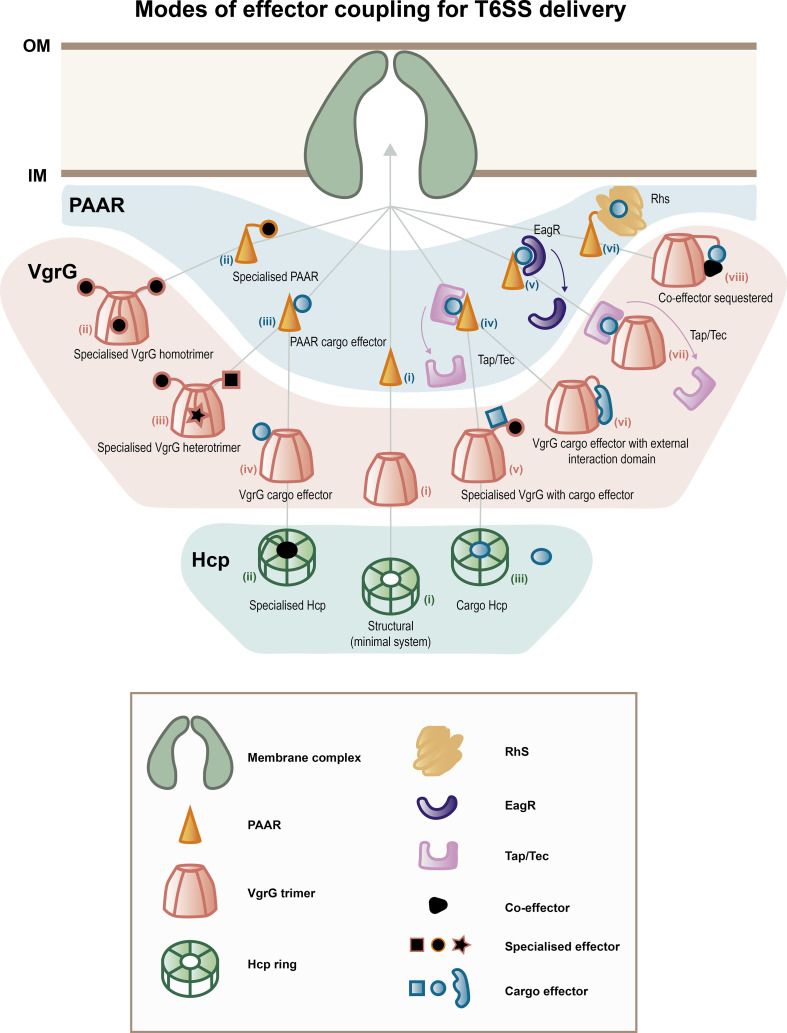
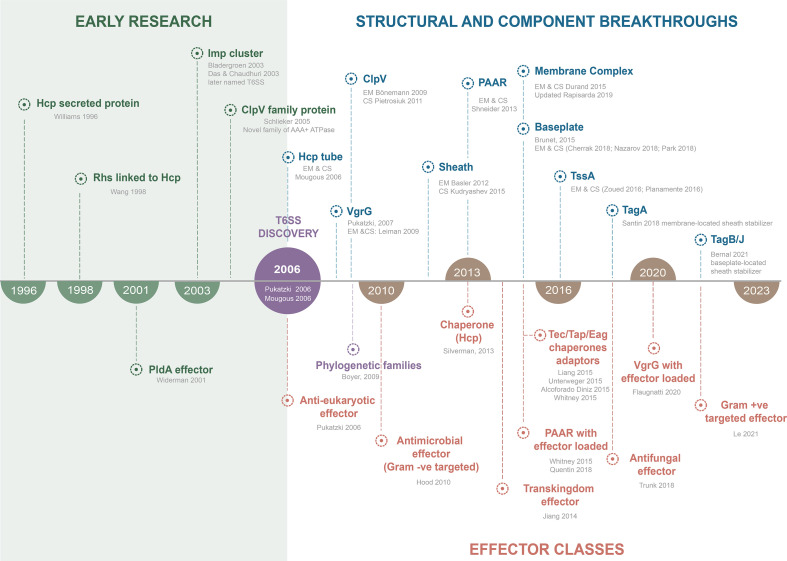
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources