

Histone bivalency regulates the timing of cerebellar granule cell development
- PMID: 37491148
- PMCID: PMC10499015
- DOI: 10.1101/gad.350594.123
Histone bivalency regulates the timing of cerebellar granule cell development
Abstract
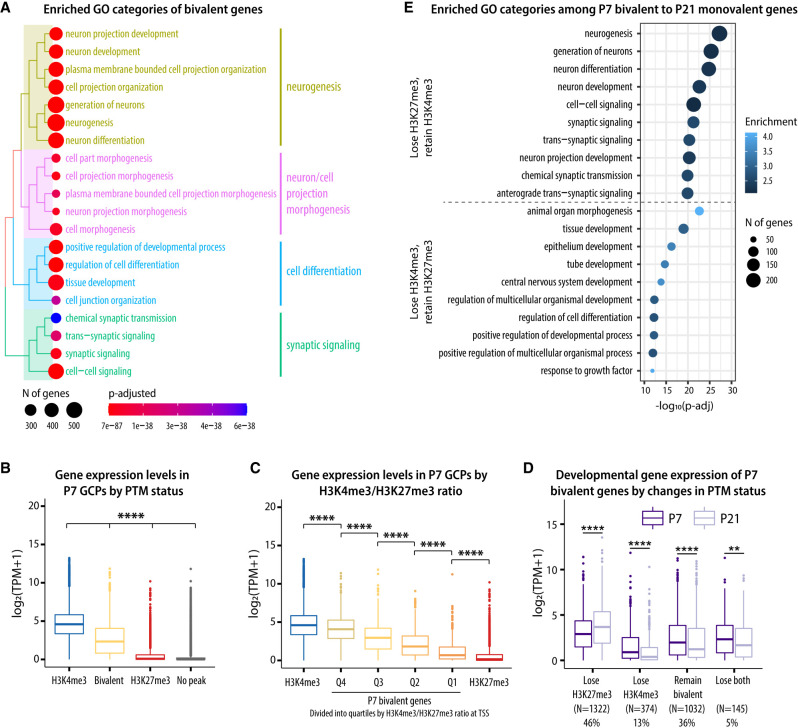
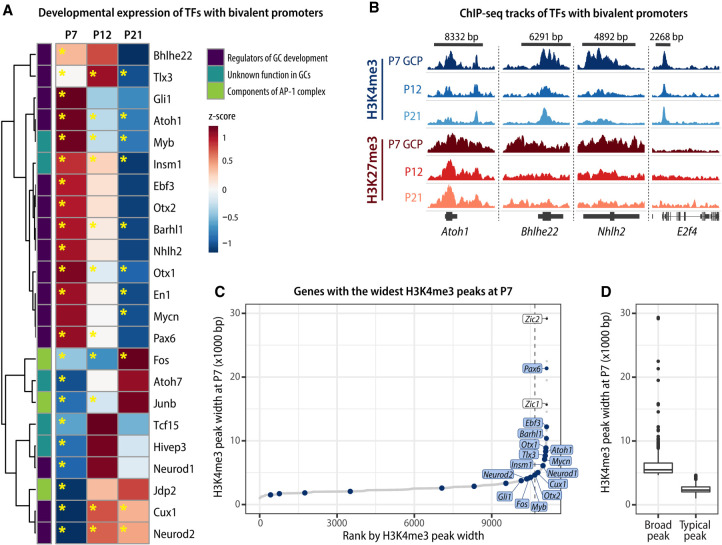
Developing neurons undergo a progression of morphological and gene expression changes as they transition from neuronal progenitors to mature neurons. Here we used RNA-seq and H3K4me3 and H3K27me3 ChIP-seq to analyze how chromatin modifications control gene expression in a specific type of CNS neuron: the mouse cerebellar granule cell (GC). We found that in proliferating GC progenitors (GCPs), H3K4me3/H3K27me3 bivalency is common at neuronal genes and undergoes dynamic changes that correlate with gene expression during migration and circuit formation. Expressing a fluorescent sensor for bivalent domains revealed subnuclear bivalent foci in proliferating GCPs. Inhibiting H3K27 methyltransferases EZH1 and EZH2 in vitro and in organotypic cerebellar slices dramatically altered the expression of bivalent genes, induced the down-regulation of migration-related genes and up-regulation of synaptic genes, inhibited glial-guided migration, and accelerated terminal differentiation. Thus, histone bivalency is required to regulate the timing of the progression from progenitor cells to mature neurons.
Keywords: H3K27me3; cerebellum; glial-guided migration; granule cells; histone bivalency; mouse.
© 2023 Mätlik et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





Update of
-
Histone bivalency regulates the timing of cerebellar granule cell development.bioRxiv [Preprint]. 2023 Feb 3:2023.02.02.526881. doi: 10.1101/2023.02.02.526881. bioRxiv. 2023. Update in: Genes Dev. 2023 Jul 1;37(13-14):570-589. doi: 10.1101/gad.350594.123. PMID: 36778390 Free PMC article. Updated. Preprint.
References
-
- Aoyama K, Oshima M, Koide S, Suzuki E, Mochizuki-Kashio M, Kato Y, Tara S, Shinoda D, Hiura N, Nakajima-Takagi Y, et al. 2018. Ezh1 targets bivalent genes to maintain self-renewing stem cells in Ezh2-insufficient myelodysplastic syndrome. iScience 9: 161–174. 10.1016/j.isci.2018.10.008 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous