

RARRES2 regulates lipid metabolic reprogramming to mediate the development of brain metastasis in triple negative breast cancer
- PMID: 37491281
- PMCID: PMC10369725
- DOI: 10.1186/s40779-023-00470-y
RARRES2 regulates lipid metabolic reprogramming to mediate the development of brain metastasis in triple negative breast cancer
Abstract
Background: Triple negative breast cancer (TNBC), the most aggressive subtype of breast cancer, is characterized by a high incidence of brain metastasis (BrM) and a poor prognosis. As the most lethal form of breast cancer, BrM remains a major clinical challenge due to its rising incidence and lack of effective treatment strategies. Recent evidence suggested a potential role of lipid metabolic reprogramming in breast cancer brain metastasis (BCBrM), but the underlying mechanisms are far from being fully elucidated.
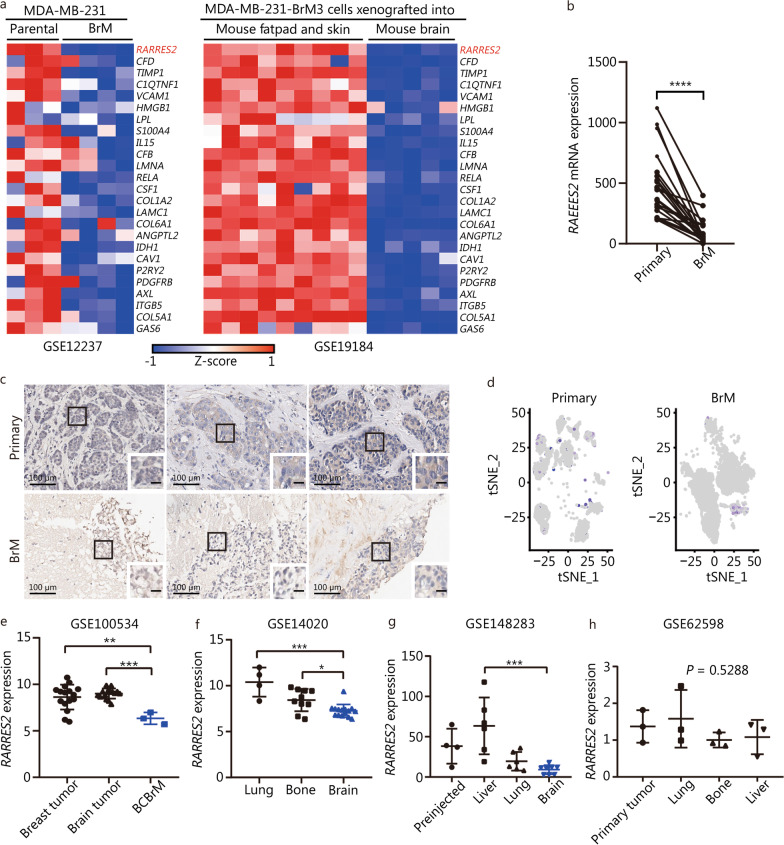
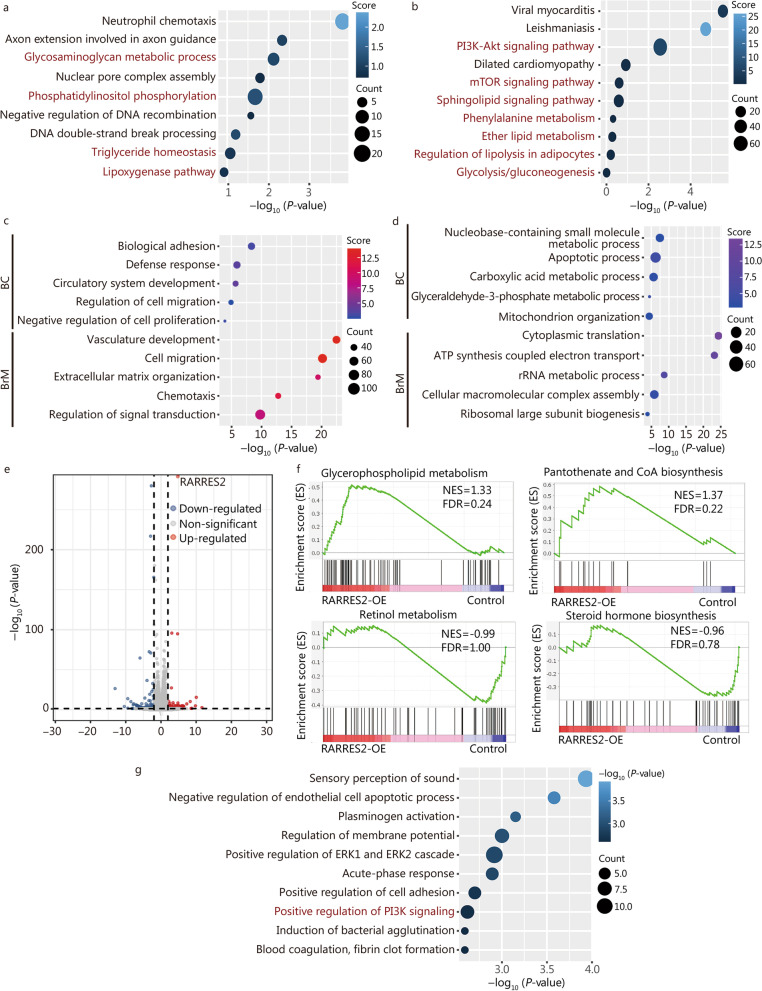
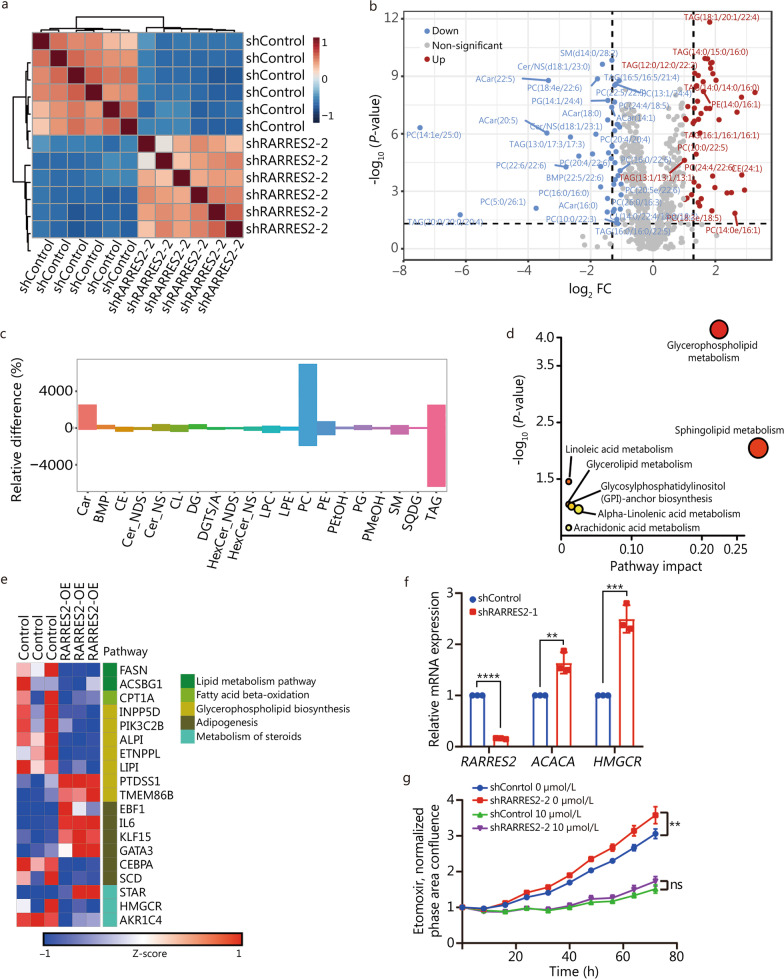
Methods: Through analysis of BCBrM transcriptome data from mice and patients, and immunohistochemical validation on patient tissues, we identified and verified the specific down-regulation of retinoic acid receptor responder 2 (RARRES2), a multifunctional adipokine and chemokine, in BrM of TNBC. We investigated the effect of aberrant RARRES2 expression of BrM in both in vitro and in vivo studies. Key signaling pathway components were evaluated using multi-omics approaches. Lipidomics were performed to elucidate the regulation of lipid metabolic reprogramming of RARRES2.
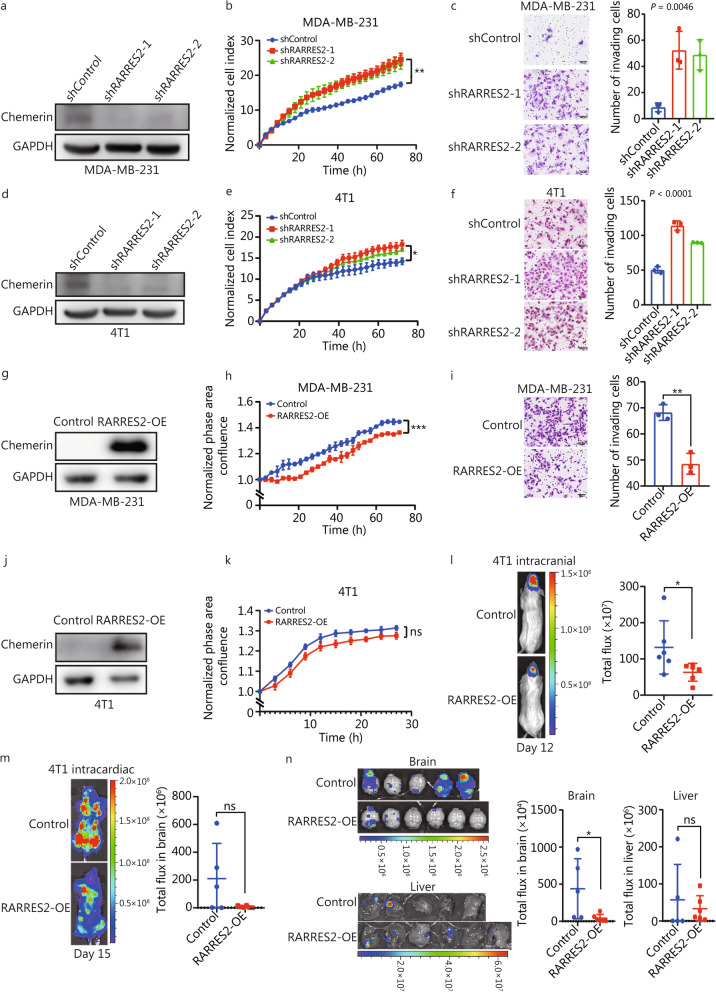
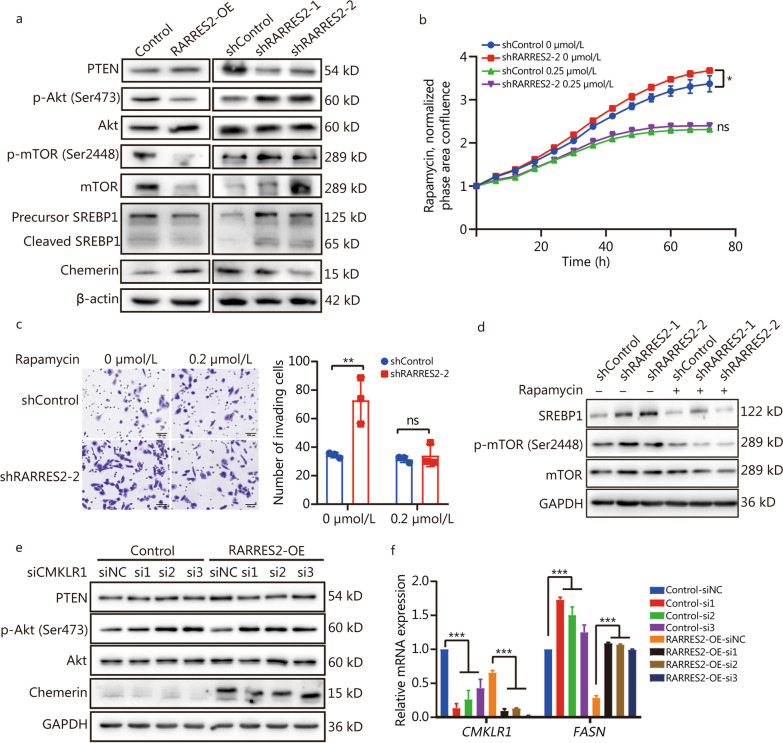
Results: We found that down-regulation of RARRES2 is specifically associated with BCBrM, and that RARRES2 deficiency promoted BCBrM through lipid metabolic reprogramming. Mechanistically, reduced expression of RARRES2 in brain metastatic potential TNBC cells resulted in increased levels of glycerophospholipid and decreased levels of triacylglycerols by regulating phosphatase and tensin homologue (PTEN)-mammalian target of rapamycin (mTOR)-sterol regulatory element-binding protein 1 (SREBP1) signaling pathway to facilitate the survival of breast cancer cells in the unique brain microenvironment.
Conclusions: Our work uncovers an essential role of RARRES2 in linking lipid metabolic reprogramming and the development of BrM. RARRES2-dependent metabolic functions may serve as potential biomarkers or therapeutic targets for BCBrM.
Keywords: Brain metastasis (BrM); Breast cancer; Lipid metabolic reprogramming; RARRES2.
© 2023. The Author(s).
Conflict of interest statement
The authors declare there is no competing interests.
Figures
Comment in
-
RARRES2's impact on lipid metabolism in triple-negative breast cancer: a pathway to brain metastasis.Mil Med Res. 2023 Sep 12;10(1):42. doi: 10.1186/s40779-023-00480-w. Mil Med Res. 2023. PMID: 37700352 Free PMC article. No abstract available.
References
-
- Lin NU, Vanderplas A, Hughes ME, Theriault RL, Edge SB, Wong YN, et al. Clinicopathologic features, patterns of recurrence, and survival among women with triple-negative breast cancer in the National Comprehensive Cancer Network. Cancer. 2012;118(22):5463–5472. doi: 10.1002/cncr.27581. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
