

Glucocorticoid dysfunction in children with severe malaria
- PMID: 37492570
- PMCID: PMC10364055
- DOI: 10.3389/fimmu.2023.1187196
Glucocorticoid dysfunction in children with severe malaria
Abstract
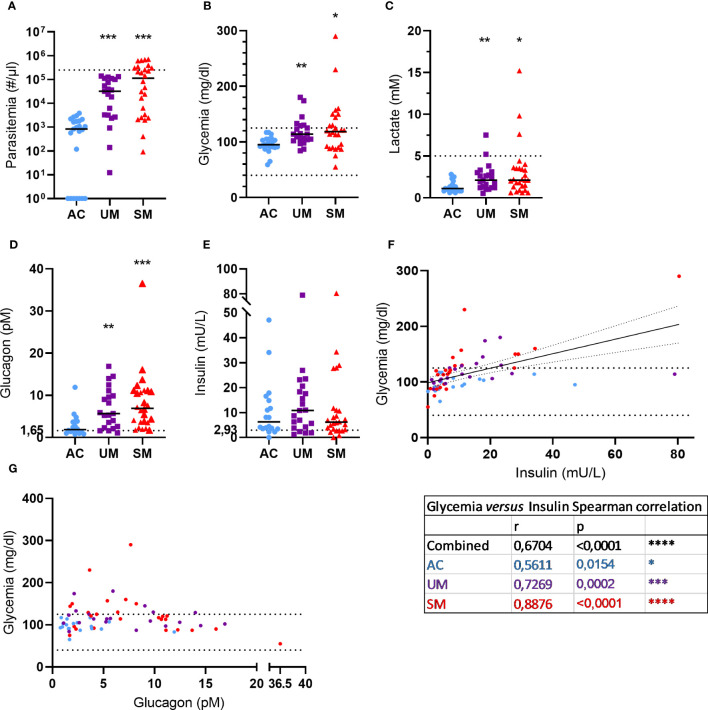
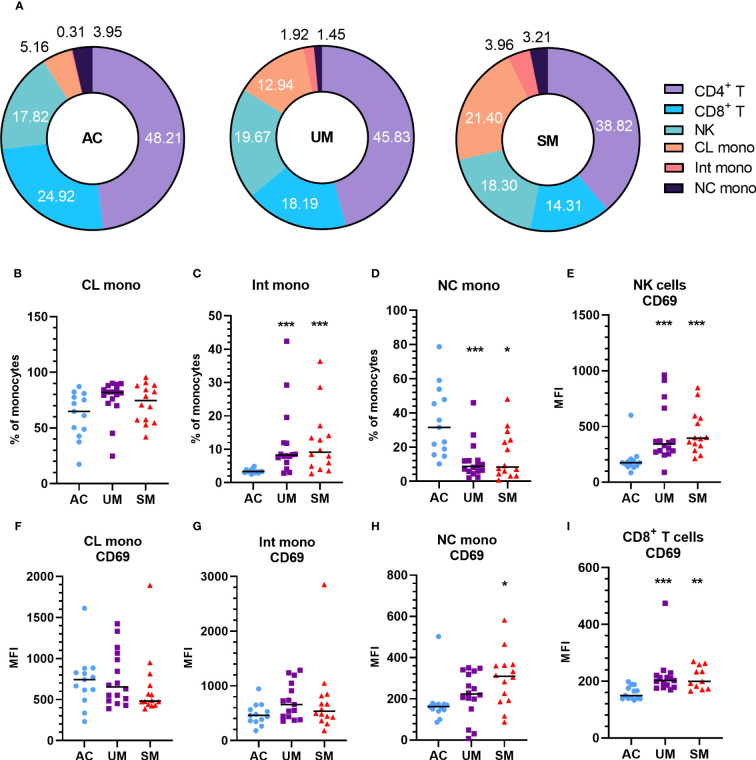
Introduction: Malaria remains a widespread health problem with a huge burden. Severe or complicated malaria is highly lethal and encompasses a variety of pathological processes, including immune activation, inflammation, and dysmetabolism. Previously, we showed that adrenal hormones, in particular glucocorticoids (GCs), play critical roles to maintain disease tolerance during Plasmodium infection in mice. Here, GC responses were studied in Cameroon in children with uncomplicated malaria (UM), severe malaria (SM) and asymptomatic controls (AC).
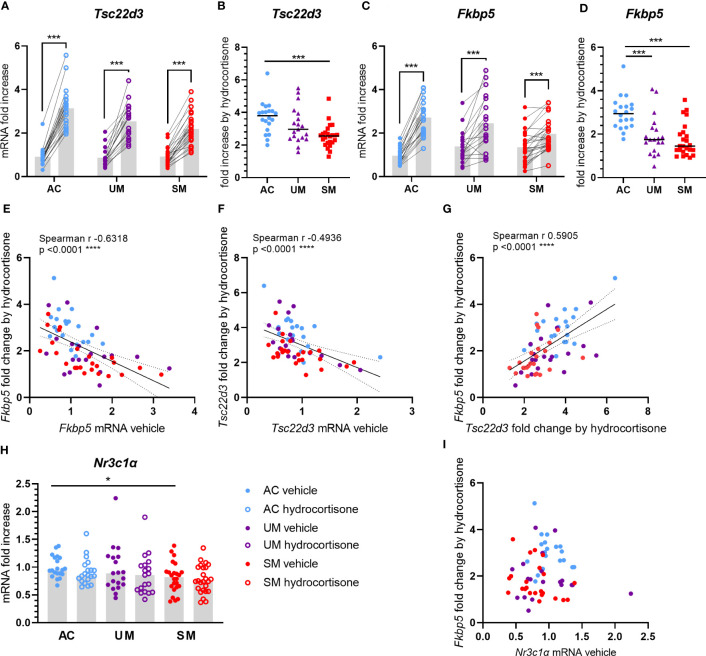
Methods: To determine the sensitivity of leukocytes to GC signaling on a transcriptional level, we measured the ex vivo induction of glucocorticoid induced leucine zipper (GILZ) and FK506-binding protein 5 (FKBP5) by GCs in human and murine leukocytes. Targeted tracer metabolomics on peripheral blood mononuclear cells (PBMCs) was performed to detect metabolic changes induced by GCs.
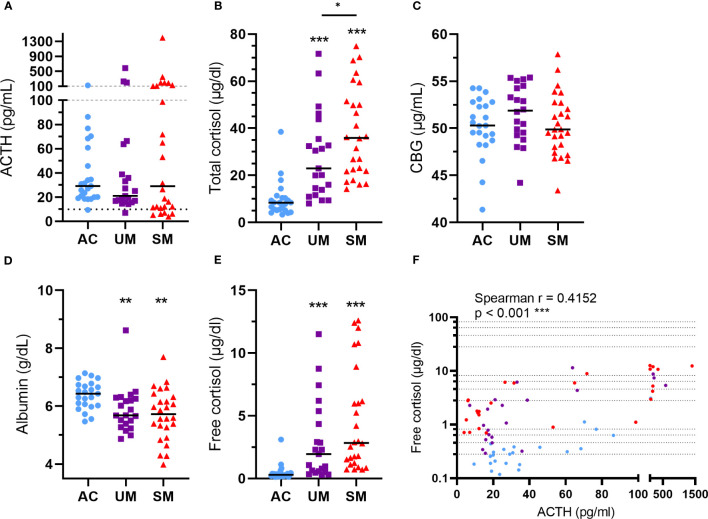
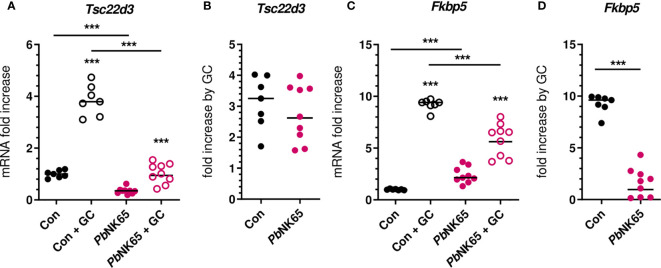
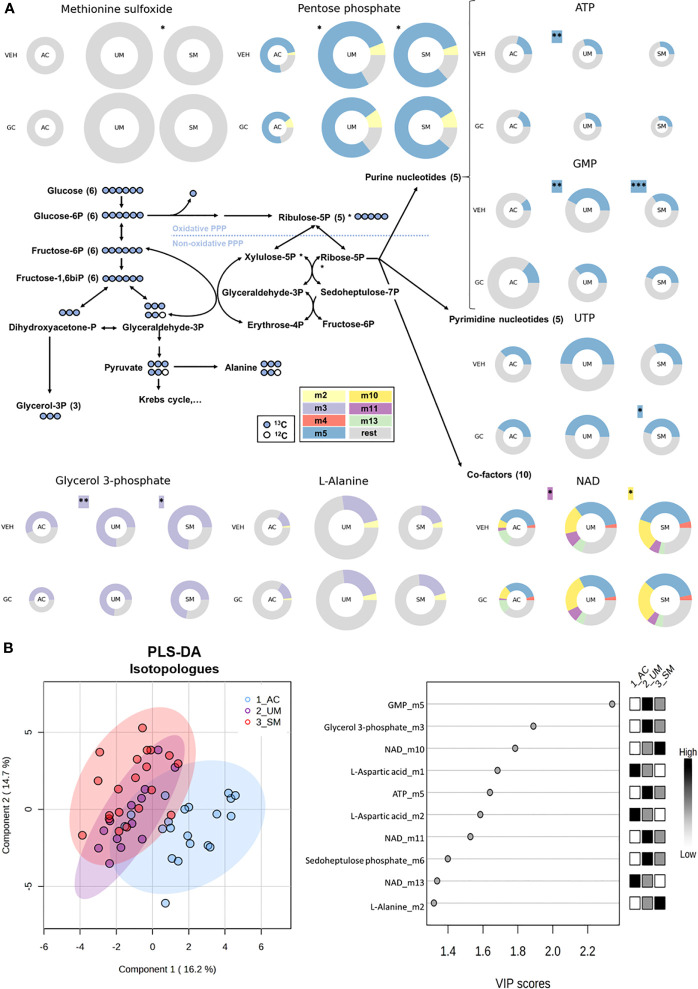
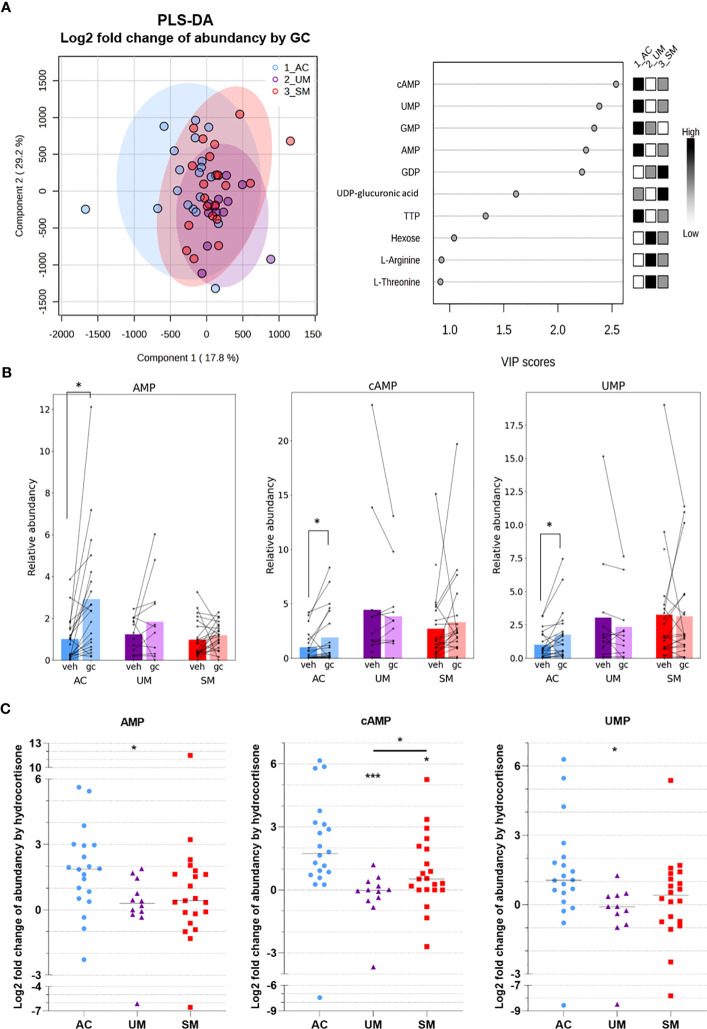
Results: Total cortisol levels increased in patients with clinical malaria compared to AC and were higher in the SM versus UM group, while cortisol binding globulin levels were unchanged and adrenocorticotropic hormone (ACTH) levels were heterogeneous. Induction of both GILZ and FKBP5 by GCs was significantly reduced in patients with clinical malaria compared to AC and in malaria-infected mice compared to uninfected controls. Increased activity in the pentose phosphate pathway was found in the patients, but this was not affected by ex vivo stimulation with physiological levels of hydrocortisone. Interestingly, hydrocortisone induced increased levels of cAMP in AC, but not in clinical malaria patients.
Discussion: Altogether, this study shows that patients with SM have increased cortisol levels, but also a decreased sensitivity to GCs, which may clearly contribute to the severity of disease.
Keywords: cortisol; glucocorticoids; malaria; metabolomics; plasmodium.
Copyright © 2023 Vandermosten, Prenen, Fogang, Dagneau de Richecour, Knoops, Donkeu, Nguefack, Taguebue, Ndombo, Ghesquière, Ayong and Van den Steen.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Insufficient glucocorticoid receptor signaling and flattened salivary cortisol profile are associated with metabolic and inflammatory indices in type 2 diabetes.J Endocrinol Invest. 2021 Jan;44(1):37-48. doi: 10.1007/s40618-020-01260-2. Epub 2020 May 11. J Endocrinol Invest. 2021. PMID: 32394161
-
Glucocorticoid receptor response and glucocorticoid-induced leucine zipper expression in neutrophils of critically ill patients with traumatic and non-traumatic brain injury.Front Endocrinol (Lausanne). 2024 Sep 9;15:1414785. doi: 10.3389/fendo.2024.1414785. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 39314520 Free PMC article.
-
Differential regulation of synthetic glucocorticoids on gene expression levels of glucocorticoid-induced leucine zipper and interleukin-2.J Clin Endocrinol Metab. 2005 May;90(5):2994-3000. doi: 10.1210/jc.2004-2298. Epub 2005 Feb 22. J Clin Endocrinol Metab. 2005. PMID: 15728202
-
Glucocorticoids and Glucocorticoid-Induced-Leucine-Zipper (GILZ) in Psoriasis.Front Immunol. 2019 Sep 13;10:2220. doi: 10.3389/fimmu.2019.02220. eCollection 2019. Front Immunol. 2019. PMID: 31572404 Free PMC article. Review.
-
Variation in glucocorticoid sensitivity and the relation with obesity.Obes Rev. 2022 Mar;23(3):e13401. doi: 10.1111/obr.13401. Epub 2021 Nov 27. Obes Rev. 2022. PMID: 34837448 Free PMC article. Review.
Cited by
-
A systematic review and meta-analysis of cortisol levels in Plasmodium infections.Sci Rep. 2024 Aug 6;14(1):18162. doi: 10.1038/s41598-024-68596-0. Sci Rep. 2024. PMID: 39107355 Free PMC article.
-
JAK/STAT inhibition protects glucocorticoid receptor knockout mice from lethal malaria-induced hypoglycemia and hyperinflammation.EMBO Mol Med. 2025 Aug;17(8):2040-2070. doi: 10.1038/s44321-025-00264-w. Epub 2025 Jul 23. EMBO Mol Med. 2025. PMID: 40702267 Free PMC article.
-
Resolution of experimental malaria-associated acute respiratory distress syndrome is Alox12 independent and shows residual inflammation.Malar J. 2025 Jul 4;24(1):216. doi: 10.1186/s12936-025-05462-y. Malar J. 2025. PMID: 40615878 Free PMC article.
References
-
- WHO World malaria report 2022 Available at: http://www.who.int/malaria/en/
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous