

Modeling epigenetic lesions that cause gliomas
- PMID: 37494934
- PMCID: PMC10530192
- DOI: 10.1016/j.cell.2023.06.022
Modeling epigenetic lesions that cause gliomas
Abstract
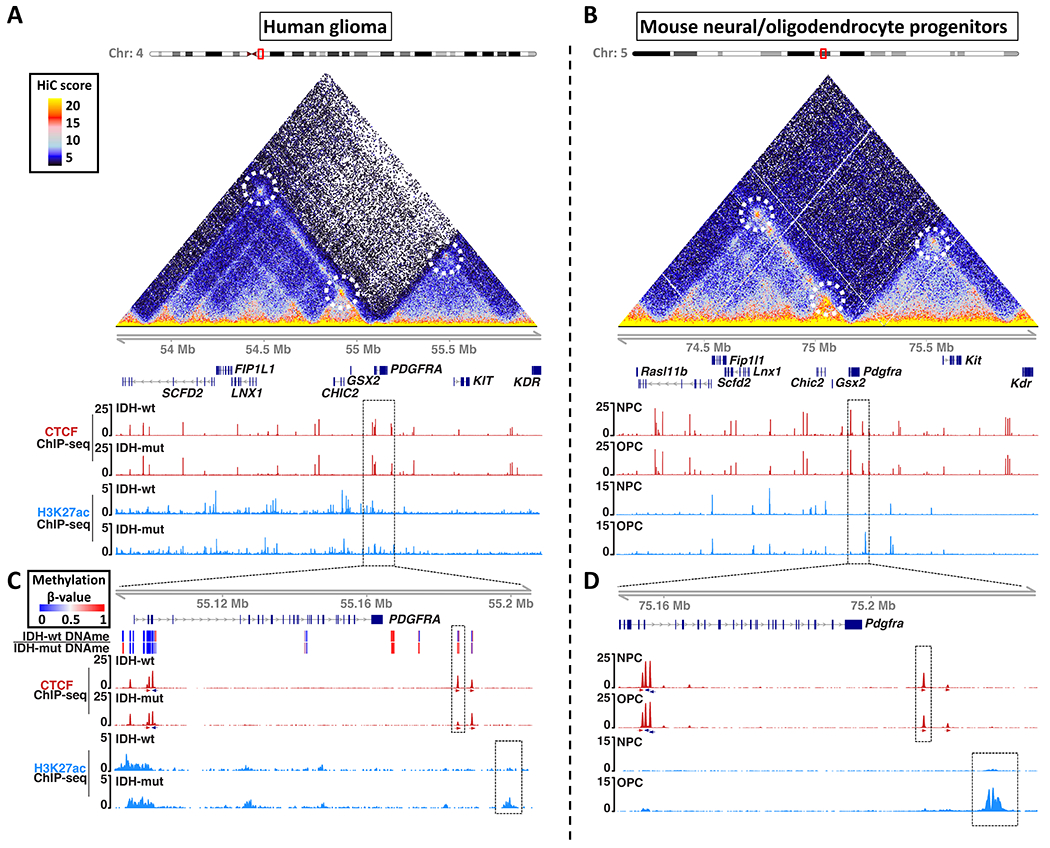
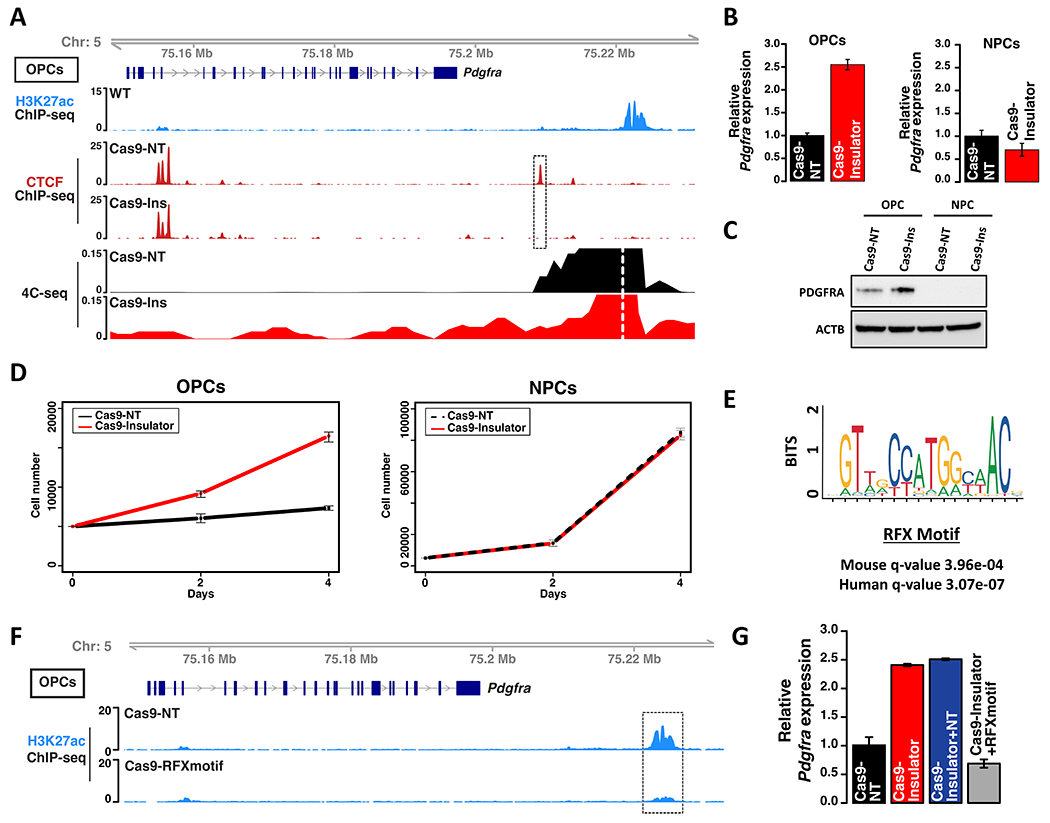
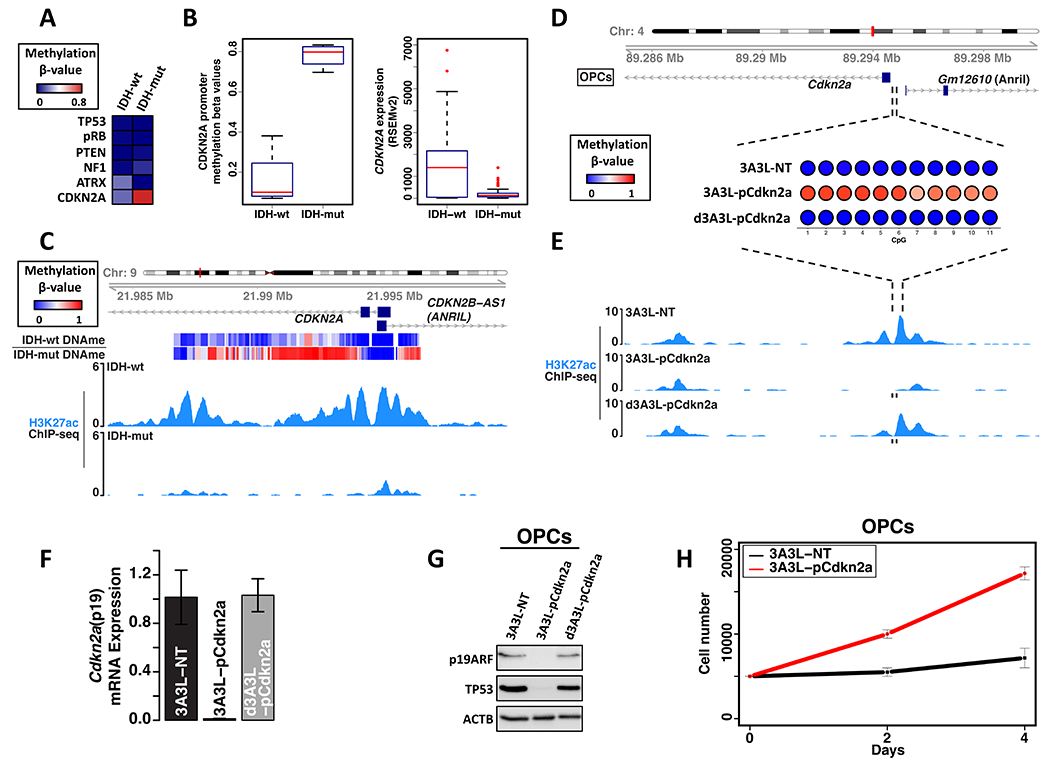
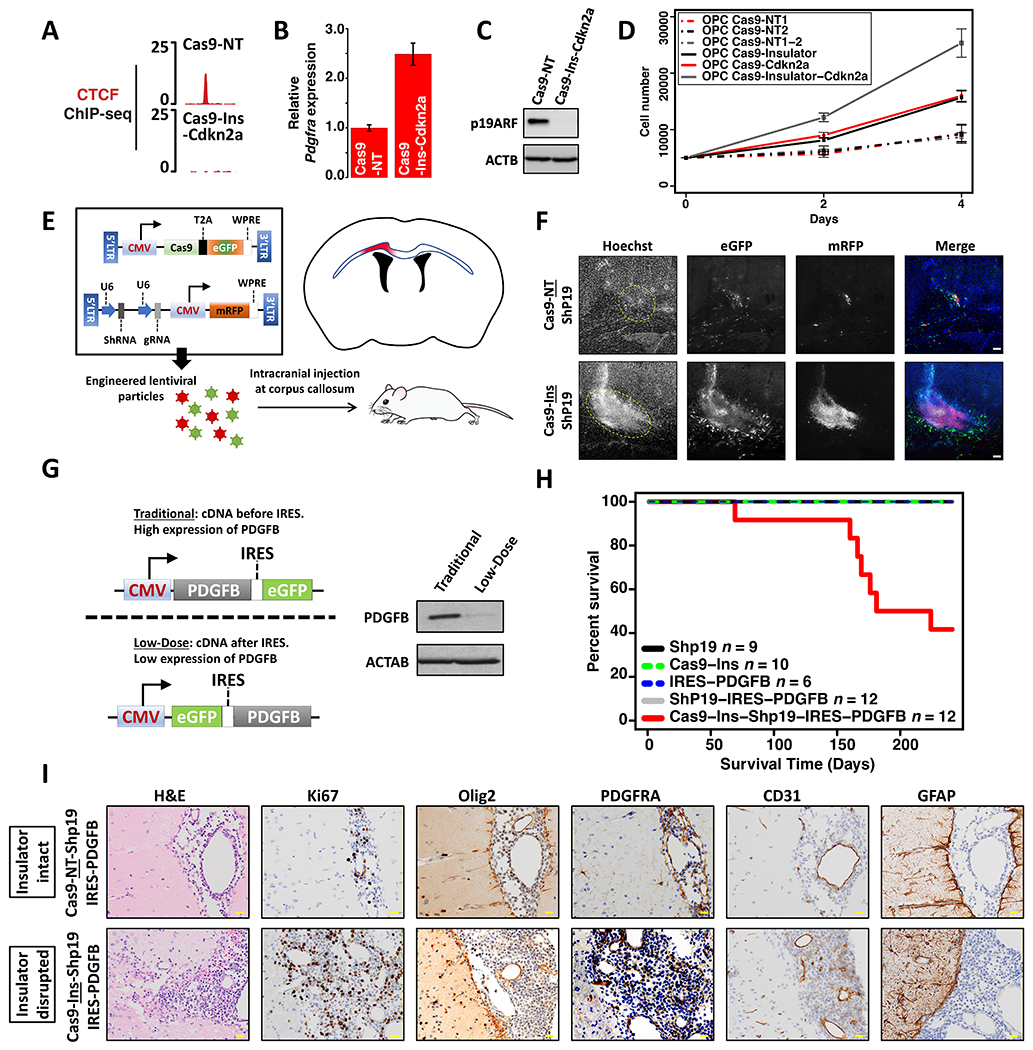
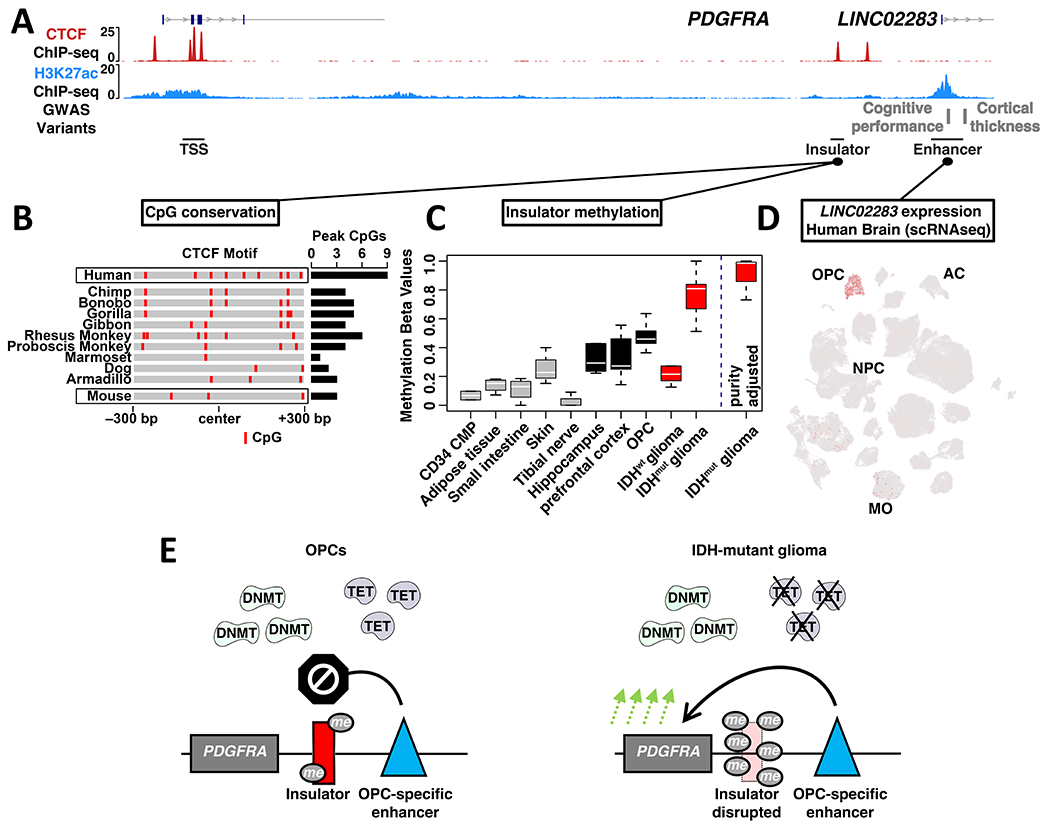
Epigenetic lesions that disrupt regulatory elements represent potential cancer drivers. However, we lack experimental models for validating their tumorigenic impact. Here, we model aberrations arising in isocitrate dehydrogenase-mutant gliomas, which exhibit DNA hypermethylation. We focus on a CTCF insulator near the PDGFRA oncogene that is recurrently disrupted by methylation in these tumors. We demonstrate that disruption of the syntenic insulator in mouse oligodendrocyte progenitor cells (OPCs) allows an OPC-specific enhancer to contact and induce Pdgfra, thereby increasing proliferation. We show that a second lesion, methylation-dependent silencing of the Cdkn2a tumor suppressor, cooperates with insulator loss in OPCs. Coordinate inactivation of the Pdgfra insulator and Cdkn2a drives gliomagenesis in vivo. Despite locus synteny, the insulator is CpG-rich only in humans, a feature that may confer human glioma risk but complicates mouse modeling. Our study demonstrates the capacity of recurrent epigenetic lesions to drive OPC proliferation in vitro and gliomagenesis in vivo.
Keywords: CDKN2A; DNA methylation; IDH mutation; PDGFRA; cell of origin; chromatin; genome topology; glioma; nuclear architecture; oligodendrocyte progenitor cells.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.E.B. discloses financial interests in Fulcrum Therapeutics, HiFiBio, Arsenal Biosciences, Chroma Medicine, Cell Signaling Technologies, and Design Pharmaceuticals.
Figures
Comment in
-
Epigenetic lesions drive gliomagenesis.Nat Rev Cancer. 2023 Oct;23(10):653. doi: 10.1038/s41568-023-00619-w. Nat Rev Cancer. 2023. PMID: 37670179 No abstract available.
References
-
- Cancer Genome Atlas Research Network, Brat DJ, Verhaak RGW, Aldape KD, Yung WKA, Salama SR, Cooper LAD, Rheinbay E, Miller CR, Vitucci M, et al. (2015). Comprehensive, Integrative Genomic Analysis of Diffuse Lower-Grade Gliomas. N Engl J Med 372, 2481–2498. 10.1056/NEJMoa1402121. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
