

The nuclear lamina is required for proper development and nuclear shape distortion in tomato
- PMID: 37503569
- PMCID: PMC10540737
- DOI: 10.1093/jxb/erad294
The nuclear lamina is required for proper development and nuclear shape distortion in tomato
Abstract
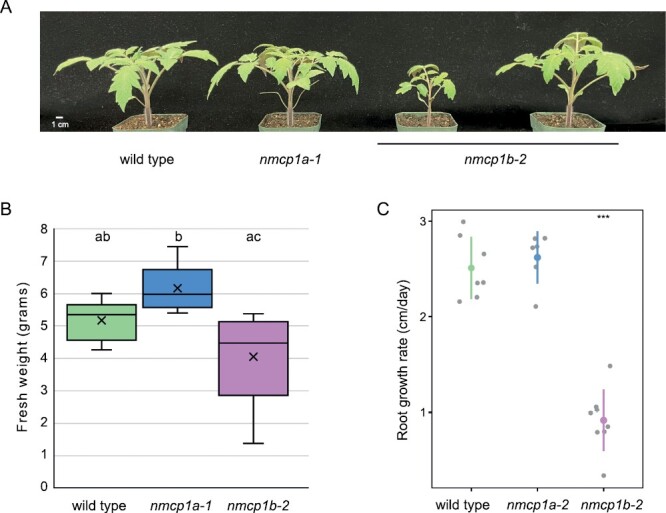
The nuclear lamina in plant cells is composed of plant-specific proteins, including nuclear matrix constituent proteins (NMCPs), which have been postulated to be functional analogs of lamin proteins that provide structural integrity to the organelle and help stabilize the three-dimensional organization of the genome. Using genomic editing, we generated alleles for the three genes encoding NMCPs in cultivated tomato (Solanum lycopersicum) to determine if the consequences of perturbing the nuclear lamina in this crop species were similar to or distinct from those observed in the model Arabidopsis thaliana. Loss of the sole NMCP2-class protein was lethal in tomato but is tolerated in Arabidopsis. Moreover, depletion of NMCP1-type nuclear lamina proteins leads to distinct developmental phenotypes in tomato, including leaf morphology defects and reduced root growth rate (in nmcp1b mutants), compared with cognate mutants in Arabidopsis. These findings suggest that the nuclear lamina interfaces with different developmental and signaling pathways in tomato compared with Arabidopsis. At the subcellular level, however, tomato nmcp mutants resembled their Arabidopsis counterparts in displaying smaller and more spherical nuclei in differentiated cells. This result argues that the plant nuclear lamina facilitates nuclear shape distortion in response to forces exerted on the organelle within the cell.
Keywords: Leaf morphology; nuclear lamina; nuclear shape; nuclear size; root growth; tomato.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Conflict of interest statement
No conflict of interest declared.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
