

Histidine Protonation and Conformational Switching in Diphtheria Toxin Translocation Domain
- PMID: 37505680
- PMCID: PMC10467104
- DOI: 10.3390/toxins15070410
Histidine Protonation and Conformational Switching in Diphtheria Toxin Translocation Domain
Abstract
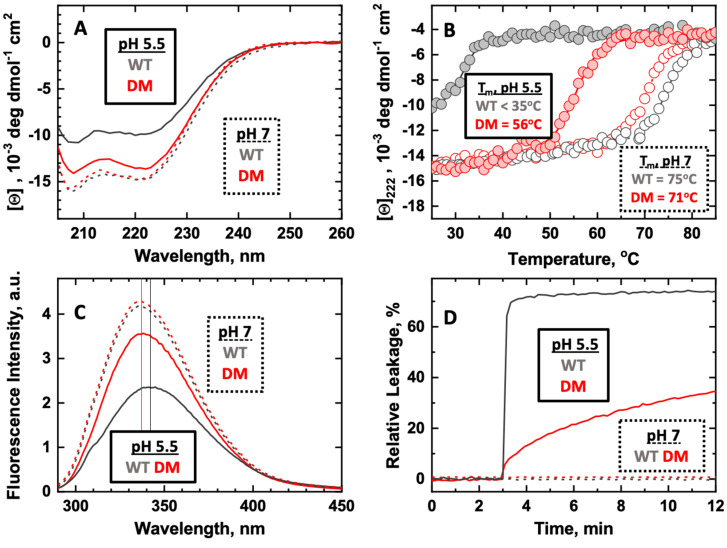
Protonation of key histidine residues has been long implicated in the acid-mediated cellular action of the diphtheria toxin translocation (T-) domain, responsible for the delivery of the catalytic domain into the cell. Here, we use a combination of computational (constant-pH Molecular Dynamics simulations) and experimental (NMR, circular dichroism, and fluorescence spectroscopy along with the X-ray crystallography) approaches to characterize the initial stages of conformational change happening in solution in the wild-type T-domain and in the H223Q/H257Q double mutant. This replacement suppresses the acid-induced transition, resulting in the retention of a more stable protein structure in solutions at pH 5.5 and, consequently, in reduced membrane-disrupting activity. Here, for the first time, we report the pKa values of the histidine residues of the T-domain, measured by NMR-monitored pH titrations. Most peaks in the histidine side chain spectral region are titrated with pKas ranging from 6.2 to 6.8. However, the two most up-field peaks display little change down to pH 6, which is a limiting pH for this protein in solution at concentrations required for NMR. These peaks are absent in the double mutant, suggesting they belong to H223 and H257. The constant-pH simulations indicate that for the T-domain in solution, the pKa values for histidine residues range from 3.0 to 6.5, with those most difficult to protonate being H251 and H257. Taken together, our experimental and computational data demonstrate that previously suggested cooperative protonation of all six histidines in the T-domain does not occur.
Keywords: NMR spectroscopy; X-ray crystallography; acidification; conformational switching; constant-pH simulations; diphtheria toxin structure; mutations of histidines.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Hoch D.H., Romero-Mira M., Ehrlich B.E., Finkelstein A., DasGupta B.R., Simpson L.L. Channels formed by botulinum, tetanus, and diphtheria toxins in planar lipid bilayers: Relevance to translocation of proteins across membranes. Proc. Natl. Acad. Sci. USA. 1985;82:1692. doi: 10.1073/pnas.82.6.1692. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
