

Deciphering the regulatory network of the NAC transcription factor FvRIF, a key regulator of strawberry (Fragaria vesca) fruit ripening
- PMID: 37506031
- PMCID: PMC10615214
- DOI: 10.1093/plcell/koad210
Deciphering the regulatory network of the NAC transcription factor FvRIF, a key regulator of strawberry (Fragaria vesca) fruit ripening
Abstract
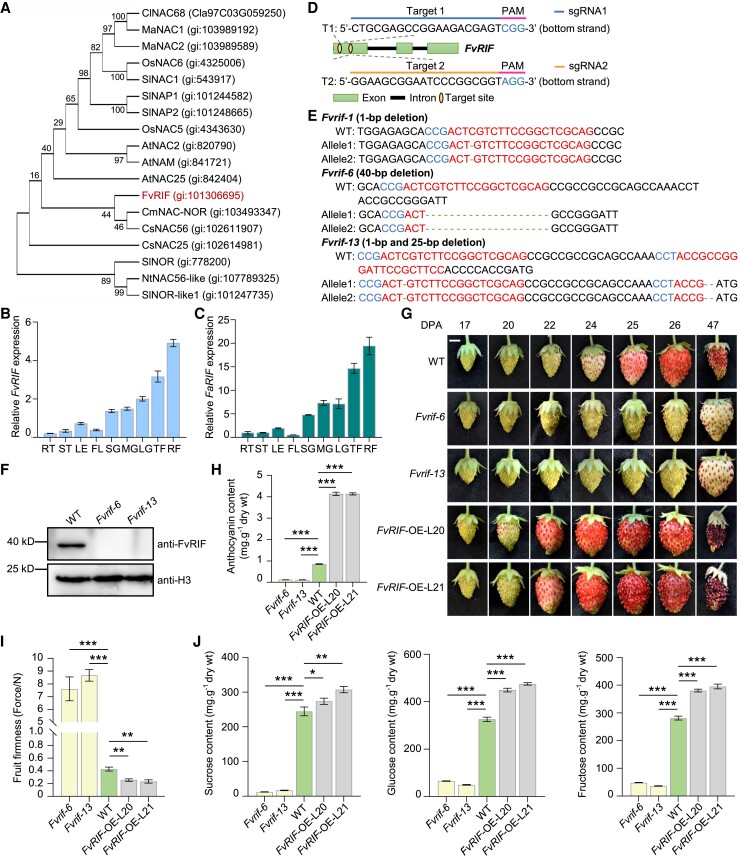
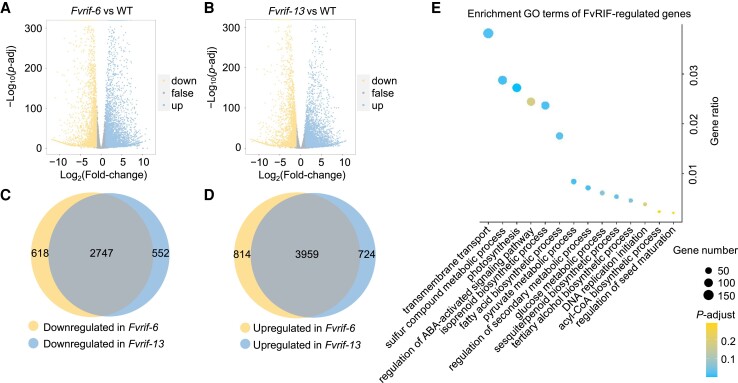
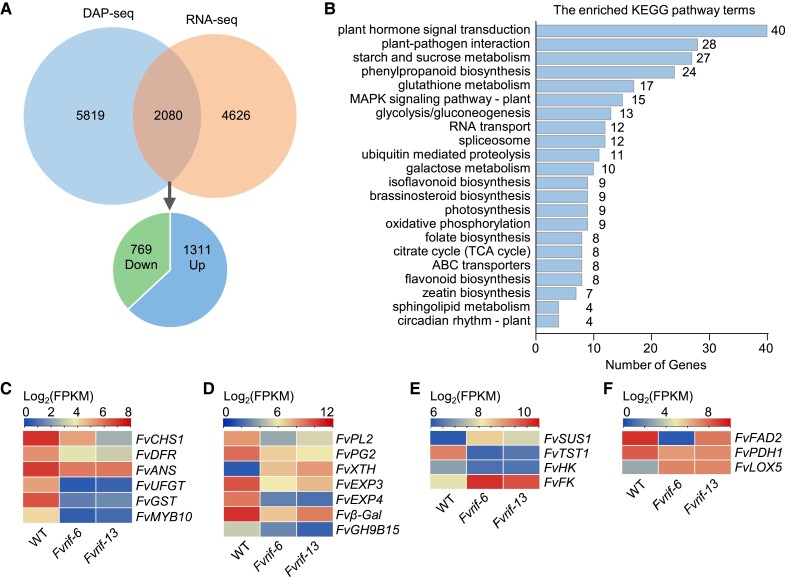
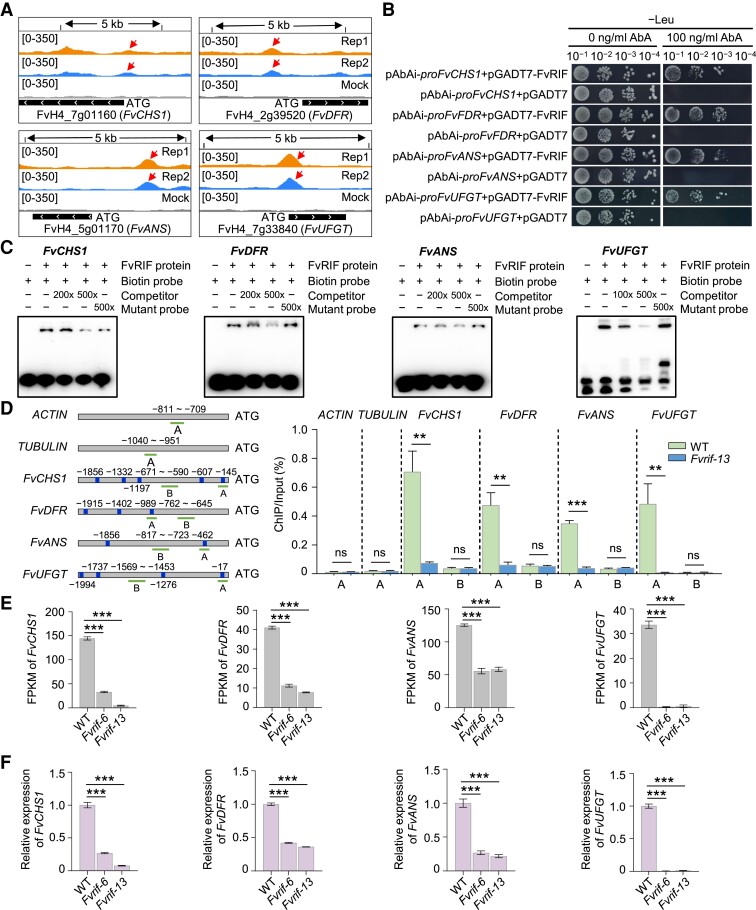
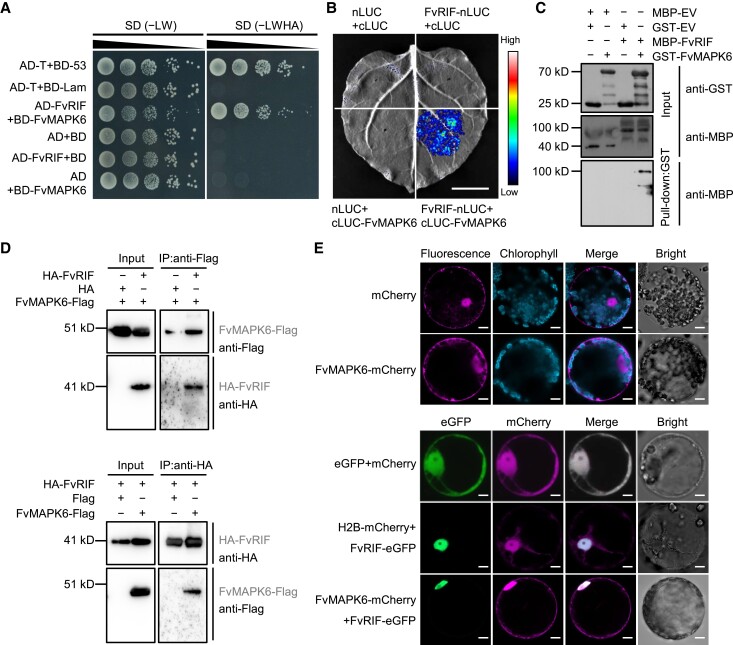
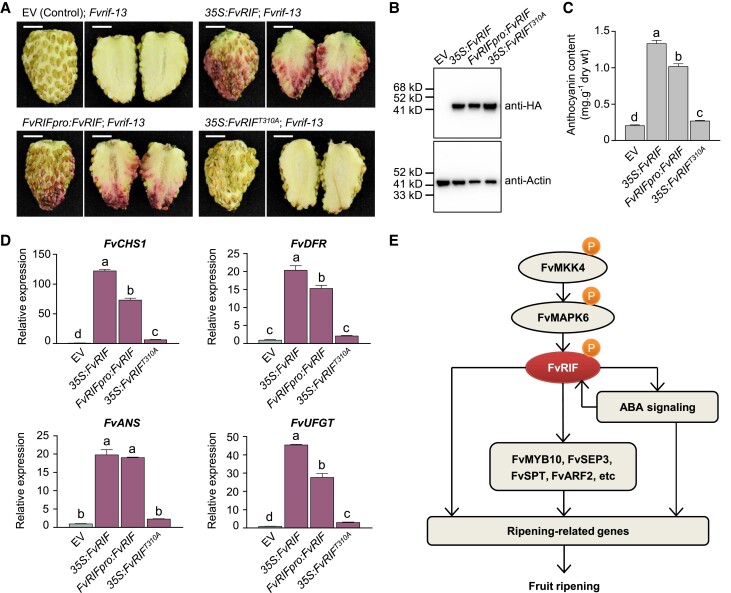
The NAC transcription factor ripening inducing factor (RIF) was previously reported to be necessary for the ripening of octoploid strawberry (Fragaria × ananassa) fruit, but the mechanistic basis of RIF-mediated transcriptional regulation and how RIF activity is modulated remains elusive. Here, we show that FvRIF in diploid strawberry, Fragaria vesca, is a key regulator in the control of fruit ripening and that knockout mutations of FvRIF result in a complete block of fruit ripening. DNA affinity purification sequencing coupled with transcriptome deep sequencing suggests that 2,080 genes are direct targets of FvRIF-mediated regulation, including those related to various aspects of fruit ripening. We provide evidence that FvRIF modulates anthocyanin biosynthesis and fruit softening by directly regulating the related core genes. Moreover, we demonstrate that FvRIF interacts with and serves as a substrate of MAP kinase 6 (FvMAPK6), which regulates the transcriptional activation function of FvRIF by phosphorylating FvRIF at Thr-310. Our findings uncover the FvRIF-mediated transcriptional regulatory network in controlling strawberry fruit ripening and highlight the physiological significance of phosphorylation modification on FvRIF activity in ripening.
© American Society of Plant Biologists 2023. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Conflict of interest statement
Conflict of interest statement. The authors declare that there is no conflict of interest.
Figures




Similar articles
-
Genome-wide analysis of the NAC transcription factor family and their expression during the development and ripening of the Fragaria × ananassa fruits.PLoS One. 2018 May 3;13(5):e0196953. doi: 10.1371/journal.pone.0196953. eCollection 2018. PLoS One. 2018. PMID: 29723301 Free PMC article.
-
Genome-wide gene network uncover temporal and spatial changes of genes in auxin homeostasis during fruit development in strawberry (F. × ananassa).BMC Plant Biol. 2024 Sep 20;24(1):876. doi: 10.1186/s12870-024-05577-5. BMC Plant Biol. 2024. PMID: 39304822 Free PMC article.
-
The FvABF3-FvALKBH10B-FvSEP3 cascade regulates fruit ripening in strawberry.Nat Commun. 2024 Dec 30;15(1):10912. doi: 10.1038/s41467-024-55294-8. Nat Commun. 2024. PMID: 39738062 Free PMC article.
-
Characterization of expansin genes and their transcriptional regulation by histone modifications in strawberry.Planta. 2021 Jul 3;254(2):21. doi: 10.1007/s00425-021-03665-6. Planta. 2021. PMID: 34216276 Review.
-
Deciphering the genetic architecture of fruit color in strawberry.J Exp Bot. 2023 Oct 31;74(20):6306-6320. doi: 10.1093/jxb/erad245. J Exp Bot. 2023. PMID: 37386925 Free PMC article. Review.
Cited by
-
LcMPK3 and LcMPK6 positively regulate fruitlet abscission in litchi.Mol Hortic. 2024 Aug 6;4(1):29. doi: 10.1186/s43897-024-00109-z. Mol Hortic. 2024. PMID: 39103914 Free PMC article.
-
Advances in genomics and genome editing for improving strawberry (Fragaria ×ananassa).Front Genet. 2024 Apr 19;15:1382445. doi: 10.3389/fgene.2024.1382445. eCollection 2024. Front Genet. 2024. PMID: 38706796 Free PMC article. Review.
-
The transcription factors AdNAC3 and AdMYB19 regulate kiwifruit ripening through brassinosteroid and ethylene signaling networks.Plant Physiol. 2025 Mar 1;197(3):kiaf084. doi: 10.1093/plphys/kiaf084. Plant Physiol. 2025. PMID: 39977254 Free PMC article.
-
Chloroplast elongation factors break the growth-immunity trade-off by simultaneously promoting yield and defence.Nat Plants. 2024 Oct;10(10):1576-1591. doi: 10.1038/s41477-024-01793-x. Epub 2024 Sep 19. Nat Plants. 2024. PMID: 39300323
-
For a Colorful Life: Recent Advances in Anthocyanin Biosynthesis during Leaf Senescence.Biology (Basel). 2024 May 9;13(5):329. doi: 10.3390/biology13050329. Biology (Basel). 2024. PMID: 38785811 Free PMC article. Review.
References
-
- Castillejo C, Waurich V, Wagner H, Ramos R, Oiza N, Munoz P, Trivino JC, Caruana J, Liu Z, Cobo N, et al. . Allelic variation of MYB10 is the major force controlling natural variation in skin and flesh color in strawberry (Fragaria spp.) fruit. Plant Cell 2020:32(12):3723–3749. 10.1105/tpc.20.00474 - DOI - PMC - PubMed
-
- Chai L, Shen Y-Y. FaABI4 is involved in strawberry fruit ripening. Sci Hortic. 2016:210(10):34–40. 10.1016/j.scienta.2016.07.015 - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
