

Structure, dynamics and transferability of the metal-dependent polyhistidine tetramerization motif TetrHis for single-chain Fv antibodies
- PMID: 37507458
- PMCID: PMC10382482
- DOI: 10.1038/s42004-023-00962-x
Structure, dynamics and transferability of the metal-dependent polyhistidine tetramerization motif TetrHis for single-chain Fv antibodies
Abstract
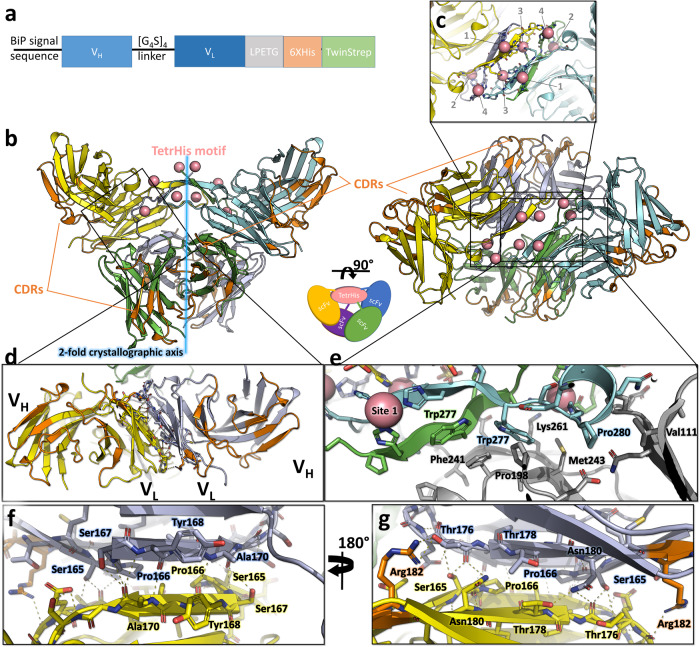
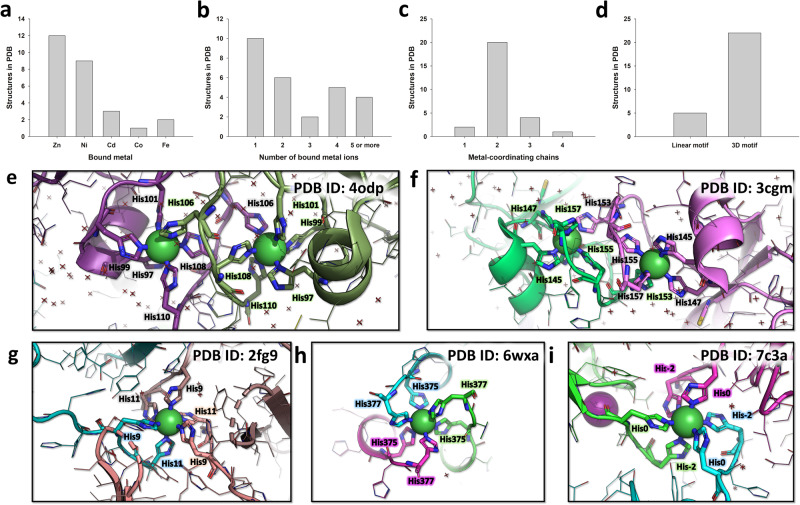
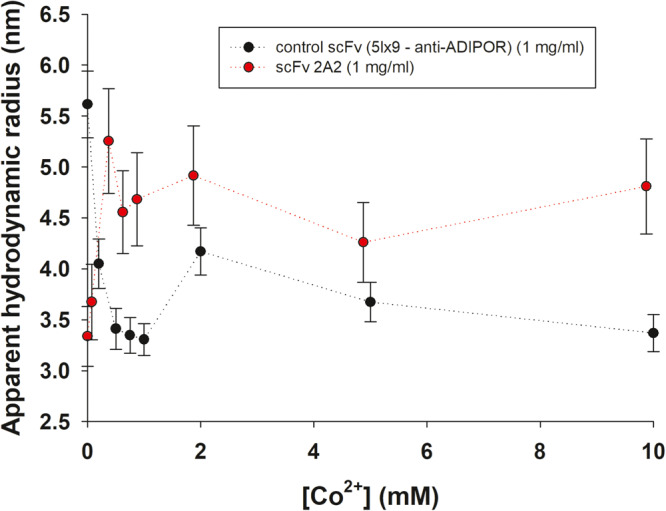
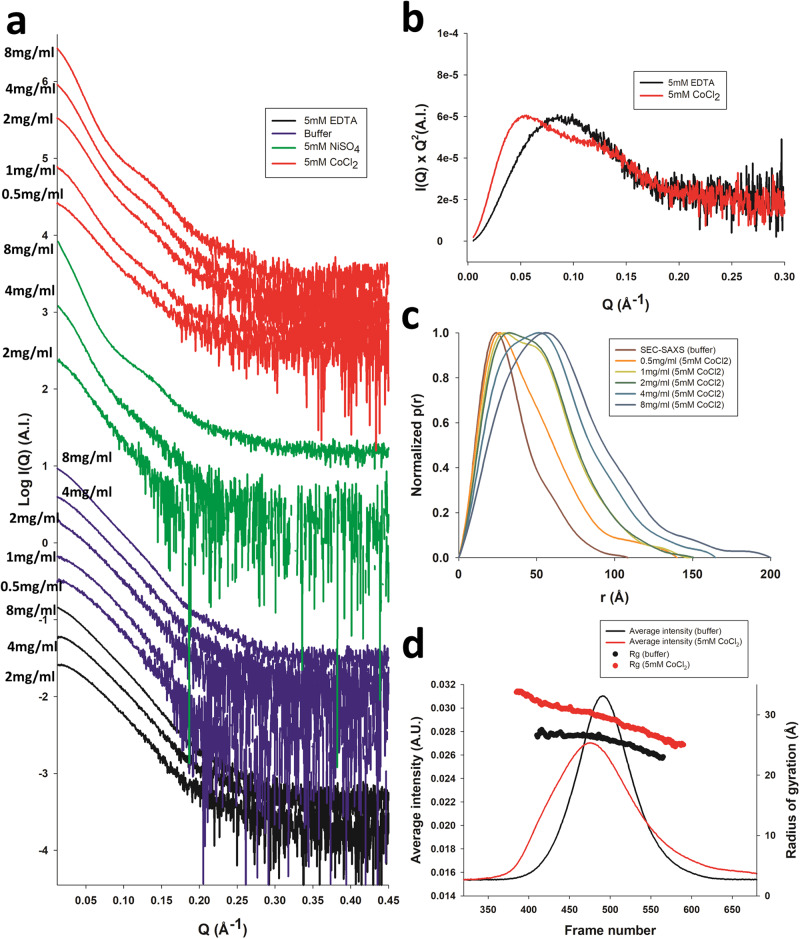
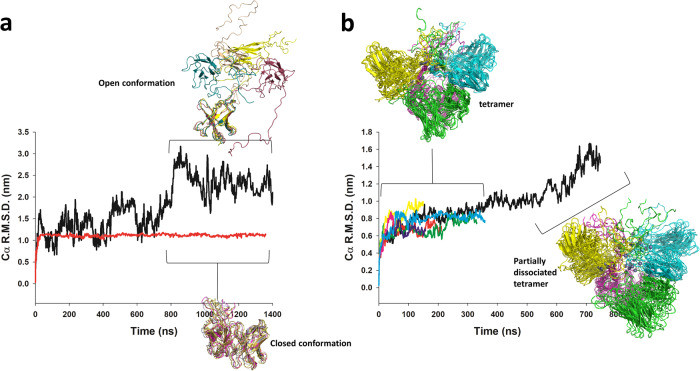
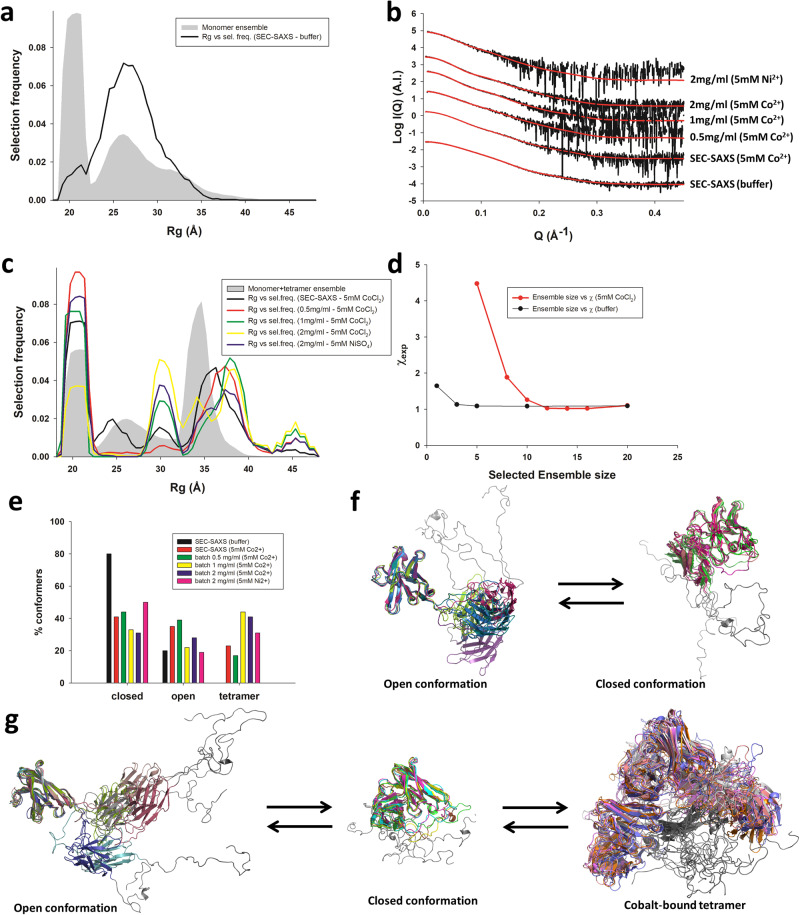
The polyhistidine (6XHis) motif is one of the most ubiquitous protein purification tags. The 6XHis motif enables the binding of tagged proteins to various metals, which can be advantageously used for purification with immobilized metal affinity chromatography. Despite its popularity, protein structures encompassing metal-bound 6XHis are rare. Here, we obtained a 2.5 Å resolution crystal structure of a single chain Fv antibody (scFv) bearing a C-terminal sortase motif, 6XHis and TwinStrep tags (LPETGHHHHHHWSHPQFEK[G3S]3WSHPQFEK). The structure, obtained in the presence of cobalt, reveals a unique tetramerization motif (TetrHis) stabilized by 8 Co2+ ions. The TetrHis motif contains four 6 residues-long β-strands, and each metal center coordinates 3 to 5 residues, including all 6XHis histidines. By combining dynamic light scattering, small angle x-ray scattering and molecular dynamics simulations, We investigated the influence of Co2+ on the conformational dynamics of scFv 2A2, observing an open/close equilibrium of the monomer and the formation of cobalt-stabilized tetramers. By using a similar scFv design, we demonstrate the transferability of the tetramerization property. This novel metal-dependent tetramerization motif might be used as a fiducial marker for cryoelectron microscopy of scFv complexes, or even provide a starting point for designing metal-loaded biomaterials.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
LinkOut - more resources
Full Text Sources
