

Inborn Errors of Purine Salvage and Catabolism
- PMID: 37512494
- PMCID: PMC10383617
- DOI: 10.3390/metabo13070787
Inborn Errors of Purine Salvage and Catabolism
Abstract
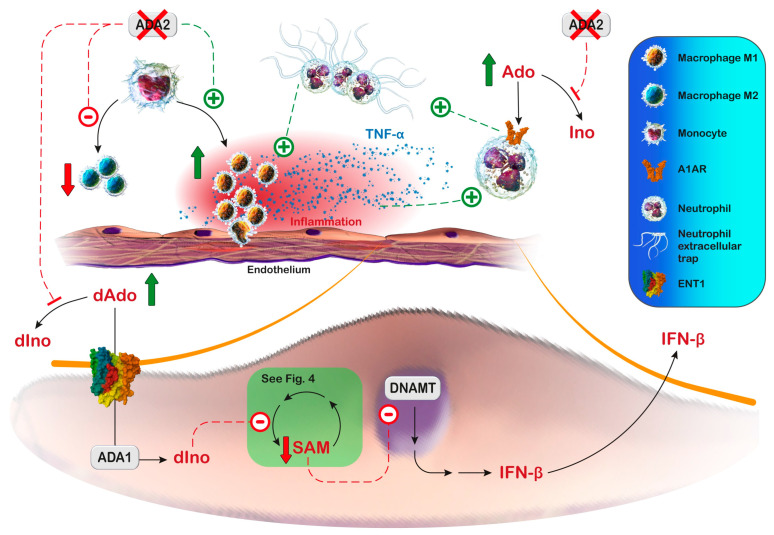
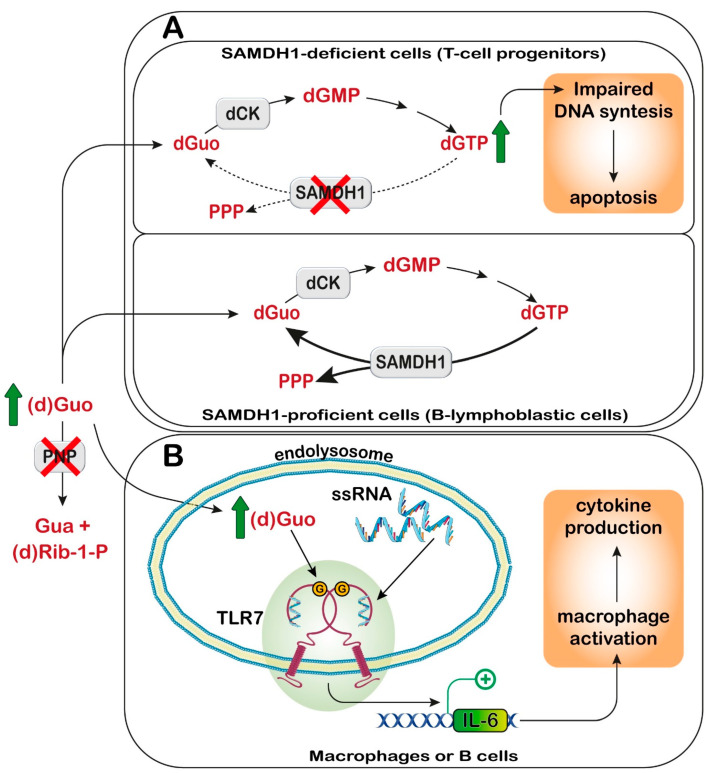
Cellular purine nucleotides derive mainly from de novo synthesis or nucleic acid turnover and, only marginally, from dietary intake. They are subjected to catabolism, eventually forming uric acid in humans, while bases and nucleosides may be converted back to nucleotides through the salvage pathways. Inborn errors of the purine salvage pathway and catabolism have been described by several researchers and are usually referred to as rare diseases. Since purine compounds play a fundamental role, it is not surprising that their dysmetabolism is accompanied by devastating symptoms. Nevertheless, some of these manifestations are unexpected and, so far, have no explanation or therapy. Herein, we describe several known inborn errors of purine metabolism, highlighting their unexplained pathological aspects. Our intent is to offer new points of view on this topic and suggest diagnostic tools that may possibly indicate to clinicians that the inborn errors of purine metabolism may not be very rare diseases after all.
Keywords: inborn errors; metabolism; metabolites; neurological disorders; neurological syndromes; purine catabolism; purine salvage; uric acid.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
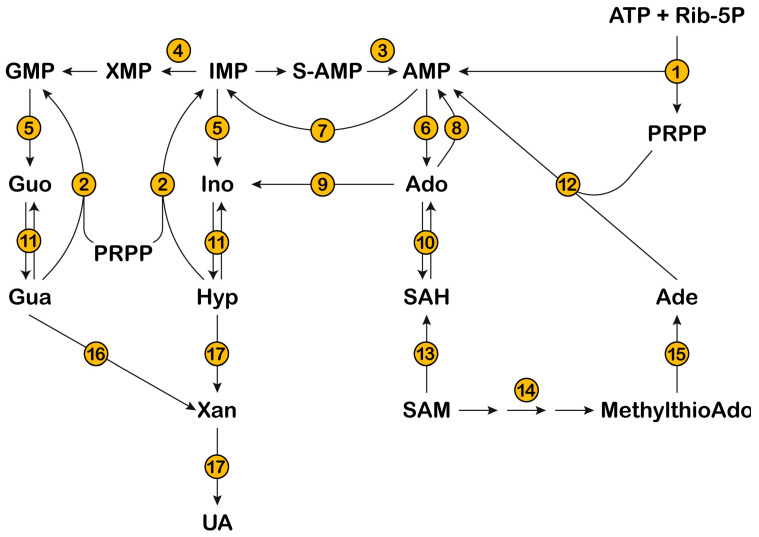
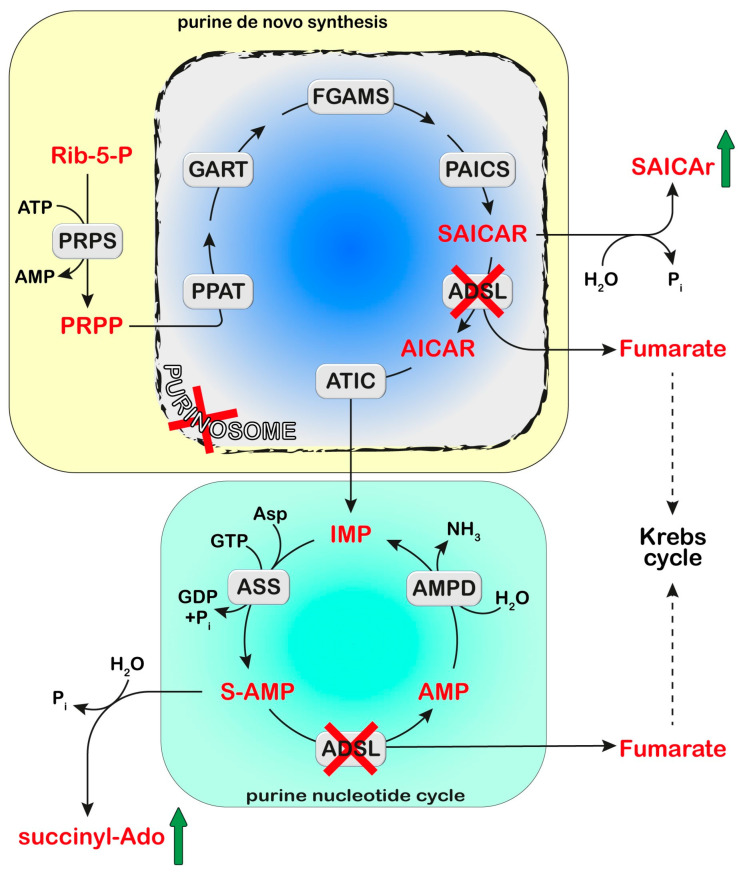
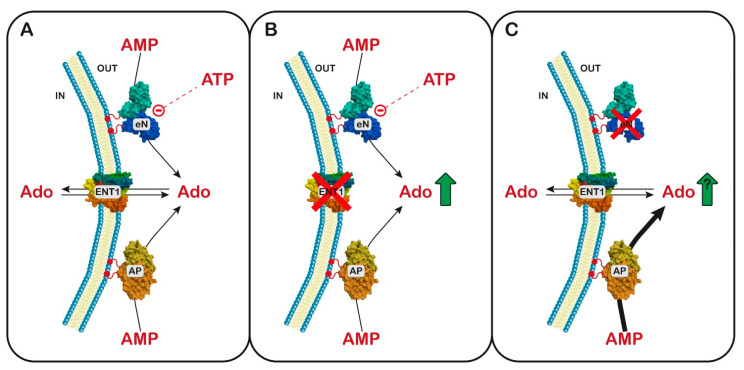
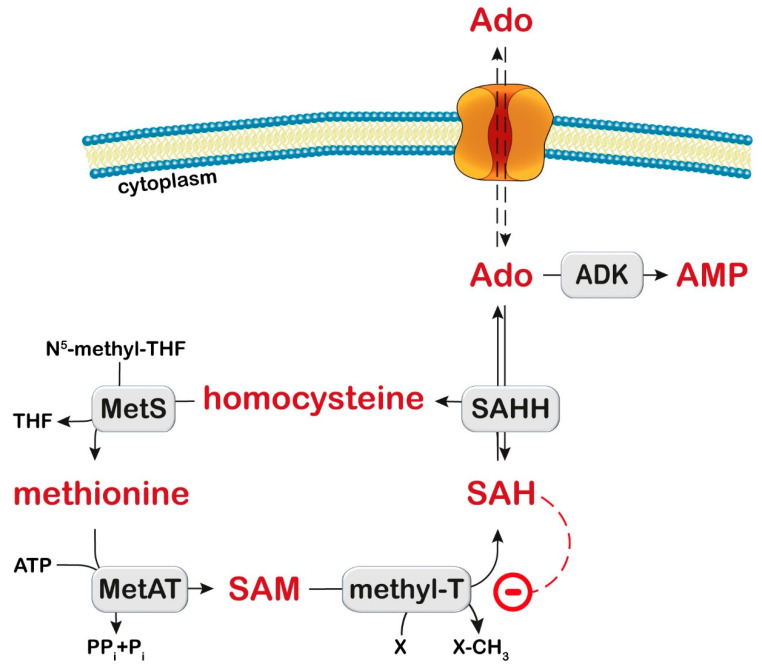
References
Publication types
LinkOut - more resources
Full Text Sources
