

Ketolysis drives CD8+ T cell effector function through effects on histone acetylation
- PMID: 37516105
- PMCID: PMC10528215
- DOI: 10.1016/j.immuni.2023.07.002
Ketolysis drives CD8+ T cell effector function through effects on histone acetylation
Abstract
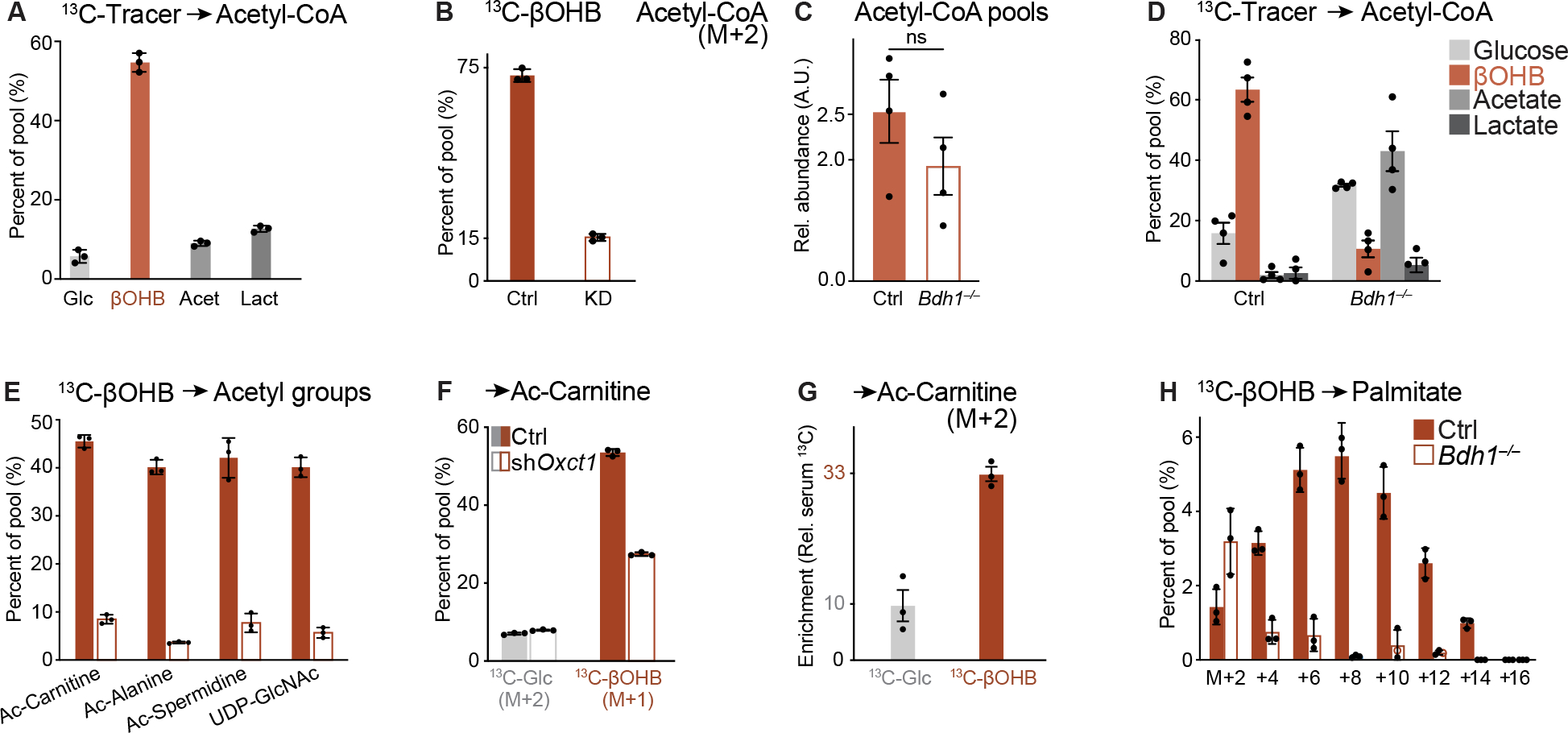
Environmental nutrient availability influences T cell metabolism, impacting T cell function and shaping immune outcomes. Here, we identified ketone bodies (KBs)-including β-hydroxybutyrate (βOHB) and acetoacetate (AcAc)-as essential fuels supporting CD8+ T cell metabolism and effector function. βOHB directly increased CD8+ T effector (Teff) cell cytokine production and cytolytic activity, and KB oxidation (ketolysis) was required for Teff cell responses to bacterial infection and tumor challenge. CD8+ Teff cells preferentially used KBs over glucose to fuel the tricarboxylic acid (TCA) cycle in vitro and in vivo. KBs directly boosted the respiratory capacity and TCA cycle-dependent metabolic pathways that fuel CD8+ T cell function. Mechanistically, βOHB was a major substrate for acetyl-CoA production in CD8+ T cells and regulated effector responses through effects on histone acetylation. Together, our results identify cell-intrinsic ketolysis as a metabolic and epigenetic driver of optimal CD8+ T cell effector responses.
Keywords: CD8(+) T cells; TCA cycle; acetyl-CoA; cancer immunology; effector function; epigenetics; ketolysis; ketone bodies; metabolism.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.G.J. is a scientific advisor for Agios Pharmaceuticals and Servier Pharmaceuticals and is a member of the Scientific Advisory Board of Immunomet Therapeutics.
Figures






References
-
- Williams MA, and Bevan MJ (2007). Effector and memory CTL differentiation. Annu. Rev. Immunol. 25, 171–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
